Fisiología de la membrana, el nervio y el músculo.
Capítulo 4: Transporte de las sustancias a través de las membranas celulares.

Las principales diferencias entre la composición de los líquidos intra- y extracelulares son las siguientes:
– Liquido extracelular: contiene concentraciones más elevadas de sodio, calcio, bicarbonato y cloruro.
– Liquido intracelular: mantiene las concentraciones de potasio, fosfatos, magnesio y proteínas más elevadas.

La membrana celular es una bicapa lipídica que constituye una barrera para el movimiento de la mayoría de las sustancias hidrosolubles, no así para las sustancias liposolubles, quienes atraviesan directamente la bicapa lipídica. Ésta membrana contiene muchas moléculas de proteínas que constituyen una vía de transporte para las sustancias hidrosolubles.

– Proteínas de los canales: son un pasaje acuoso para el movimiento de iones a través de la membrana.
– Proteínas transportadoras: a ellas se unen moléculas o iones específicos que generan cambios estructurales en dichas proteínas desplazando así las moléculas a través de la membrana.
El transporte a través de la membrana celular se produce de dos posibles formas:
– Difusión: es un movimiento molecular aleatorio a través de espacios intermoleculares. La energía que hace que se produzca la difusión es la energía del movimiento cinético normal de la materia.
– Transporte activo: movimiento de sustancias en combinación con una proteína transportadora y contra un gradiente electroquímico. Este proceso requiere de una fuente de energía adicional a la energía cinética.
~ Difusión.
Es el movimiento de las moléculas en líquidos o gases y se divide en dos subtipos:
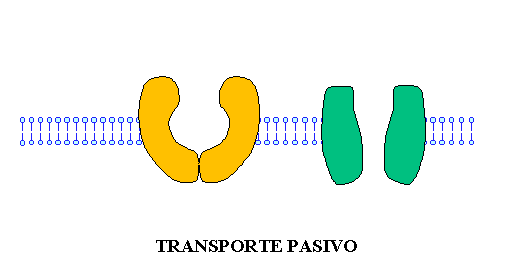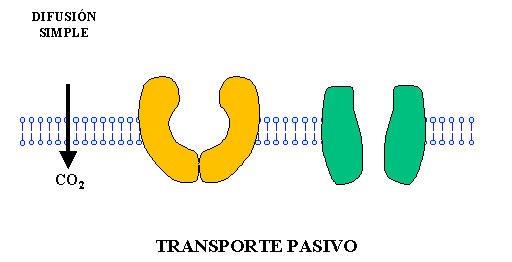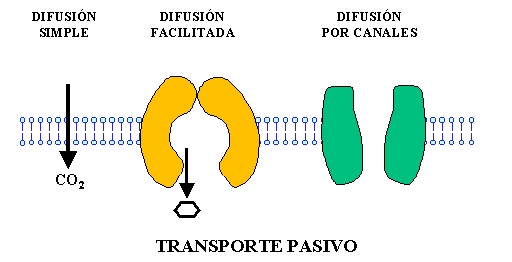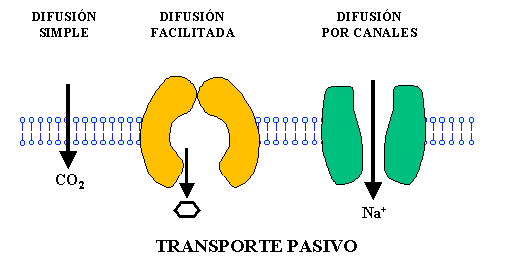
– Difusión simple: Las moléculas se mueven a través de la membrana sin unirse a proteínas transportadoras ya sea 1) a través de los intersticios de la bicapa lipídica, y 2) a través de canales proteicos.
– Difusión facilitada: Una proteína transportadora ayuda al paso de las moléculas al unirse con ellas químicamente y facilitar su desplazamiento a través de la membrana.
La velocidad con la que ocurre la difusión simple de una sustancia es directamente proporcional a su liposolubilidad. El oxígeno, nitrógeno, dióxido de carbono, gases anestésicos y la mayoría de los alcoholes tienen un grado de liposolubilidad elevada que puede difundirse directamente en la bicapa lipídica a través de a membrana celular, función que puede verse reflejada en el intercambio gaseoso de oxígeno y dióxido de carbono a nivel alveolar.

El agua y otras moléculas insolubles en lípidos, como los iones, se difunden a través de canales proteicos. Estos canales presentan una permeabilidad selectiva, consecuencia de las características del propio canal, de esta forma existe una regulación de lo que entra y sale de la célula.

La activación de los canales proteicos está controlado de los maneras:
– Activación por voltaje: La conformación molecular de la compuerta está controlada por el potencial eléctrico que se establece a través de la membrana celular.
– Activación química: Las compuertas se abren por la unión de una sustancia química a la proteína, produciendo un cambio estructural en la membrana de la proteína. este proceso se denomina activación química o activación por ligando.
La difusión facilitada también se denomina difusión mediada por un transportador debido a que las moléculas que normalmente no pueden atravesar la membrana utilizan la ayuda de una proteína transportadora específica.

La velocidad con la que se difunden las partículas en este proceso depende de la velocidad máxima con que la molécula de proteína transportadora puede sufrir cambios estructurales.
Factores que influyen en la velocidad neta de difusión:
– Permeabilidad: se expresa como la velocidad neta de difusión de la sustancia a través de cada unidad de superficie de la membrana para la diferencia de concentración.
– Diferencia de concentración: la velocidad de difusión neta es proporcional a la diferencia de concentración de la sustancia que difunde a ambos lados de la membrana.
– Potencial eléctrico: las cargas eléctricas de los iones hacen que se muevan a través de la membrana aún cuando no hay ninguna diferencia de concentración que produzca el movimiento.
Cuando la diferencia de concentración aumenta a un nivel suficientemente alto como a la diferencia del potencial eléctrico, los dos efectos se contrarrestan entre sí, creando un estado de equilibrio electroquímico.
~ Ósmosis.
Es el proceso de movimiento neto de agua causado por la diferencia de concentración de soluto.

La diferencia de presión necesaria para interrumpir la ósmosis es la presión osmótica. El factor que determina la presión osmótica de un solución es la concentración de dicha solución en función al número de partículas por unidad de volumen, pero no en función de la masa del soluto.
~ Transporte activo de sustancias a través de la membrana.
El transporte activo puede desplazar una sustancia contra un gradiente electroquímico. Un gradiente electroquímico es la suma de todas las fuerzas de difusión (diferencia de concentración, diferencia eléctrica o una diferencia de presión) que actúan en la membrana.
Se divide en dos tipos, en función de la fuente de energía utilizada. En ambos casos en transporte depende de las proteínas de membrana.
– Transporte activo primario: La energía se obtiene directamente de la división del ATP en ADP Y P o de algún otro fosfato de alta energía.
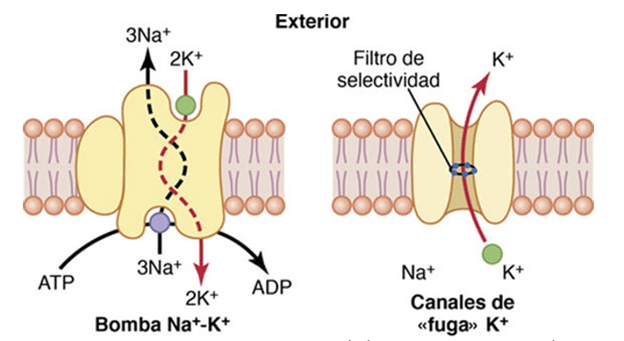
Un ejemplo significativo de transporte activo primario es la Bomba sodio – potasio la cuál transporta 3 moléculas de sodio hacia el exterior de la célula por cada 2 moléculas de potasio bombeadas hacia el interior de la célula.

– Transporte activo secundario: en éste, la energía procede de la energía almacenada en forma de diferencias de concentración iónica entre los dos lados de la membrana que se originó principalmente del transporte activo primario.
El cotransporte y contratransporte son dos tipos de transporte activo secundario.


Capítulo 5: Potenciales de membrana y potenciales de acción.

Hay potenciales eléctricos a través de las membranas de prácticamente todas las células del cuerpo. Algunas células como las nerviosas y musculares, tienen la particularidad de ser excitables, lo que significa que son capaces de generar impulsos electroquímicos en sus membranas.
Física básica de los potenciales de membrana.
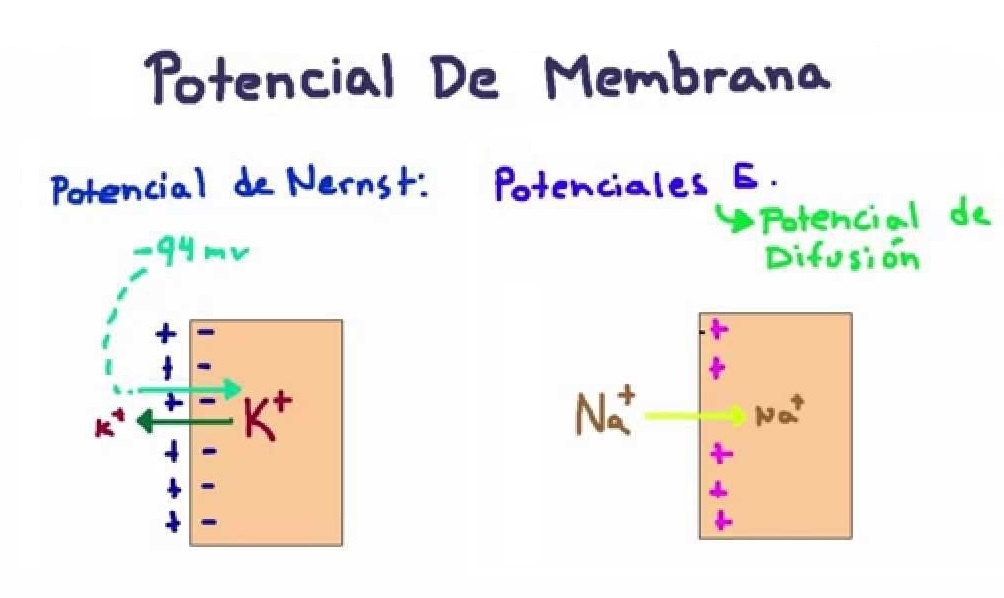
Una diferencia de concentraciones de iones puede producir un potencial de membrana.
– Potencial de difusión del potasio: La membrana celular de las neuronas es altamente permeable a los iones potasio, quienes tienden a difundir hacia el exterior a favor de su gradiente de concentración. Como los iones potasio tienen carga positiva, la pérdida de iones potasio desde la célula crea un potencial negativo en su interior el cual es lo suficientemente grande como para bloquear la difusión neta de potasio posterior.
– Potencial de difusión de sodio: Los iones sodio se difundirían hacía la célula por la alta concentración de sodio que hay en el exterior, esto crearía un potencial positivo dentro de ella, y en cuestión de milisegundos, el potencial de membrana aumentaría lo suficiente como para bloquear la difusión neta de sodio al interior de la célula.
El potencial de membrana que se opone a la difusión neta de un ion a través de la membrana se denomina : Potencial de Nernst.
La ecuación de Nernst describe la relación del potencial de difusión con la diferencia de concentración y se escribe de la siguiente manera:
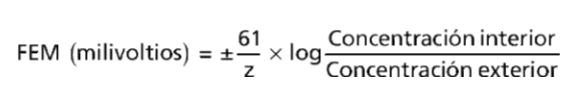
Cuando se utiliza esta fórmula habitualmente se asume que el potencial del líquido extracelular que está fuera de la membrana se mantiene a un nivel de potencial cero, y que el potencial de Nernst es el potencial que está en el interior de la membrana.
La ecuación de Goldman se utiliza para calcular el potencial de difusión cuando la membrana es permeable a varios iones diferentes:
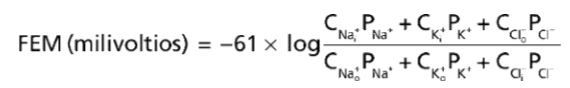
A partir de la ecuación de Goldman se hacen evidentes varios puntos clave. En primer lugar, los iones sodio, potasio y cloruro son los iones más importantes que participan en la generación de los potenciales de membrana en las fibras nerviosas y musculares, así como en las células neuronales del sistema nervioso. El gradiente de concentración de cada uno de estos iones a través de la membrana ayuda a determinar el voltaje del potencial de membrana.
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90 mV . Es decir, el potencial en el interior de la fibra es 90 mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma.
La bomba Na+-K+ también genera grandes gradientes de concentración para el sodio y el potasio a través de la membrana nerviosa en reposo.
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega a su extremo.
Las sucesivas fases del potencial de acción son las siguientes.
Fase de reposo
La fase de reposo es el potencial de membrana en reposo antes del comienzo del potencial de acción. Se dice que la membrana está «polarizada» durante esta fase debido al potencial de membrana negativo de –90 mV que está presente.
Fase de despolarización
En este momento la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado «polarizado» normal de –90 mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva, un proceso denominado despolarización.
Fase de repolarización
En un plazo de algunas diezmilésimas de segundo después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal, que se denomina repolarización de la membrana.
Capitulo 6 y 7 : Contracción del músculo esquelético.

Aproximadamente el 40% del cuerpo es músculo esquelético, y tal vez otro 10% es músculo liso y cardíaco.
Todos los músculos esqueléticos están formados por numerosas fibras cuyo diámetro varía entre 10 y 80 μm. Cada una de estas fibras está formada por subunidades cada vez más pequeñas.
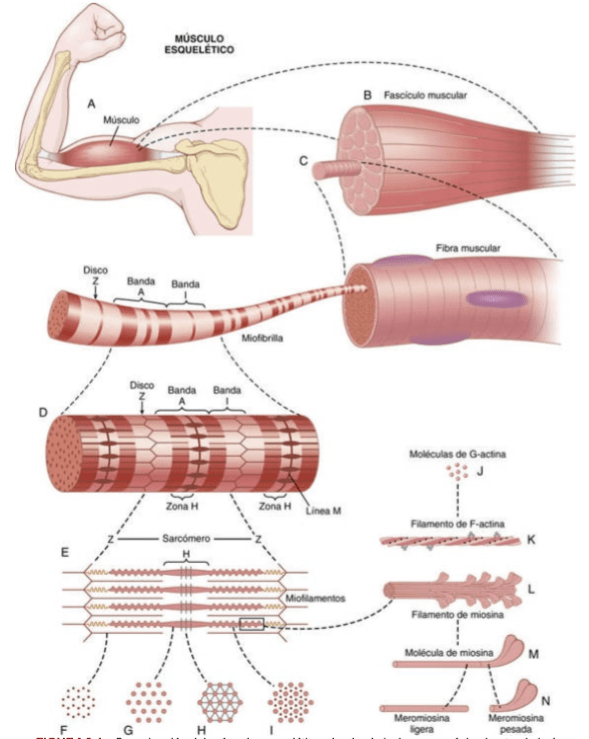
En la mayoría de los músculos esqueléticos, las fibras se extienden a lo largo de toda la longitud del músculo. Todas las fibras, excepto alrededor de un 2%, habitualmente están inervadas por una sola terminación nerviosa localizada cerca del punto medio de la fibra.
Las miofibrillas están formadas por filamentos de actina y miosina. Cada fibra muscular está formada por cientos de miofibrillas que a su vez está formada por filamentos de actina y miosina adyacentes entre sí.

El sarcoplasma es el fluido intracelular entre las miofibrillas.
Las muchas miofibrillas de cada fibra muscular están yuxtapuestas suspendidas en la fibra muscular. Los espacios entre las miofibrillas están llenos de un líquido intracelular denominado sarcoplasma, que contiene grandes cantidades de potasio, magnesio y fosfato, además de múltiples enzimas proteicas.
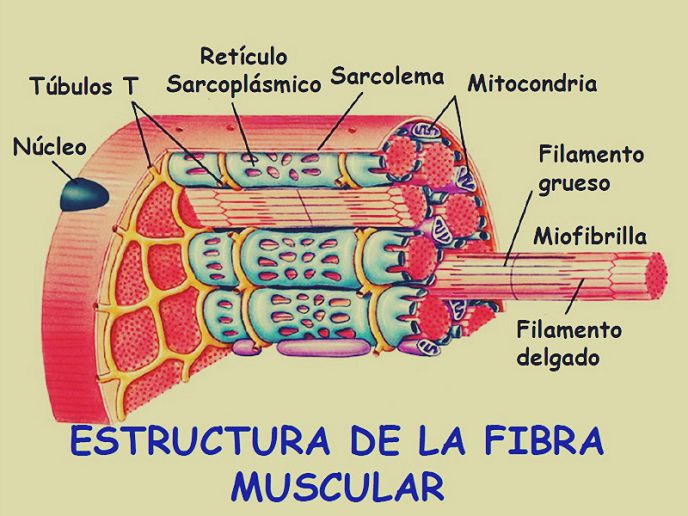
Hay muchas mitocondrias que están dispuestas paralelas a las miofibrillas. Estas mitocondrias proporcionan a las miofibrillas en contracción grandes cantidades de energía en forma de trifosfato de adenosina (ATP), que es formado por las mitocondrias.
El retículo sarcoplásmico es un retículo endoplásmico especializado de músculo esquelético. Este retículo tiene una organización especial que es muy importante para regular el almacenamiento, la liberación y la recaptación de calcio y, por tanto, para controlar la contracción muscular.
Mecanismo general de la contracción muscular.
El inicio y la ejecución de la contracción muscular se producen en las siguientes etapas secuenciales:
1. Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares.

2. En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.

3. La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membrana.
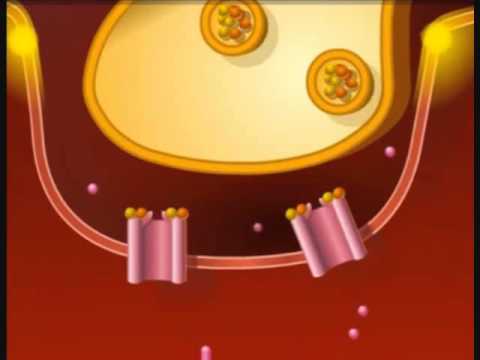
4. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.

5. El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
6. El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo.

7. Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.

8. Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.

Capitulo 8 : Excitación y contracción del músculo liso.
El músculo liso está formado por fibras de 1 a 5 um de diámetro y de sólo 20 a 500 um de longitud. La contracción se da por las mismas fuerzas de atracción entre los filamentos de miosina y actina.

Tipos de músculo liso
- El músculo liso se distingue por:
- Dimensiones físicas
- Organización en fascículos o láminas Respuesta a diferentes tipos de estímulos
- Características de la inervación
- Función
En general se puede dividir en dos tipos principales:
- Músculo liso multiunitario

Formado por fibras musculares lisas separadas y discretas. Cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa. La superficie externa de estas fibras está cubierta por una capa delgada de sustancia similar a una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí, pero la característica más importante es que cada una de las fibras se puede contraer independientemente de las demás, y su control se ejerce principalmente por señales nerviosas.
- Músculo liso unitario o monounitario

Masa de de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad. Las fibras habitualmente están dispuestas en láminas o fascículos, y sus membranas celulares están adheridas entre sí en múltiples puntos. Las membranas celulares están unidas por muchas uniones en hendidura a través de las cuales los iones pueden fluir libremente desde una célula muscular a otra y hacer que las fibras musculares se contraigan simultáneamente.

También se conoce como músculo liso sincitial debido a sus interconexiones sincitiales entre las fibras y como músculo liso visceral porque se encuentra en la pared de la mayor parte de las vísceras del cuerpo.

Mecanismo contráctil en el músculo liso
Base química de la contracción del músculo liso
El músculo liso contiene filamentos tanto de actina como de miosina, pero no contiene el complejo de troponina normal.
El proceso contráctil es activado por los iones calcio, y el trifosfato de adenosina (ATP) se degrada a difosfato de adenosina (ADP) para proporcionar la energía para la contracción.

Base física de la contracción del músculo liso
El músculo liso no tiene la misma disposición estriada de los filamentos de actina y miosina. Grandes números de filamentos de actina unidos a los denominados cuerpos densos, los cuales tienen la misma función que los discos Z. Algunos de estos cuerpos están unidos a la membrana celular y otros están dispersos en el interior de la célula. Algunos de los cuerpos densos de la membrana de células adyacentes están unidos entre sí por puentes proteicos intercelulares. La fuerza de contracción se transmite de unas células a otras a través de estos enlaces. Interpuestos entre los filamentos de actina de la fibra muscular están los filamentos de miosina.
La mayor parte de los filamentos de miosina tiene lo que se denomina puentes cruzados lateropolares, dispuestos de tal manera que los puentes de un lado basculan en una dirección y los del otro lado basculan en la dirección opuesta. Esto permite que la miosina tire de un filamento de actina en una dirección en un lado a la vez que simultáneamente tira de otro filamento de actina en la dirección opuesta en el otro lado, esto permite que las células musculares lisas se contraigan hasta el 80% de su longitud.
Comparación de la contracción del músculo liso con la contracción del músculo estriado
La mayor parte de las contracciones del músculo liso son contracciones tónicas prolongadas, que a veces duran horas o incluso días.
- Ciclado lento de los puentes cruzados de miosina
La rapidez del ciclado de los puentes transversos de miosina en el músculo liso es más lenta. La fracción de tiempo que los puentes cruzados permanecen unidos a los filamentos de actina está muy aumentada en el músculo liso. Una posible razón del ciclado lento es que las cabezas de los puentes cruzados tienen una actividad ATPasa mucho menor.
- Baja necesidad de energía para mantener la contracción del músculo liso
Para mantener la misma tensión de contracción en el músculo liso es necesario de 1/10 a 1/300 de energía. Sólo es necesaria una molécula de ATP para cada ciclo. La baja utilización de energía por el músculo liso es muy importante para la economía energética global del cuerpo.
- Lentitud del inicio de la contracción y relajación del tejido muscular liso total
Un tejido muscular liso típico comienza a contraerse de 50 a 100 ms después de ser excitado, alcanza la contracción completa aproximadamente 0,5 s después, y después la fuerza contráctil disminuye en 1 a 2 segundos más, dando un tiempo total de contracción de 1 a 3 s. La contracción de algunos tipos puede ser tan corta como 0,2 s o tan larga como 30 s. El inicio de la contracciónes es mucho más lento en respuesta a los iones calcio.
- La fuerza máxima de contracción muscular es a menudo mayor en el músculo liso que en el músculo esquelético
La fuerza de contracción del músculo liso es mayor, hasta 4 a 6 kg/cm2 de área transversal. Esto se debe al período prolongado de unión de los puentes cruzados de miosina a los filamentos de actina.
- El mecanismo de «cerrojo» facilita el mantenimiento prolongado de las contracciones del músculo liso

Una vez que el músculo liso ha generado la contracción máxima, la magnitud de la excitación continuada habitualmente se puede reducir a mucho menos del nivel inicial, a pesar de lo cual el músculo mantiene su fuerza de contracción completa. Además, la energía que se consume para mantener la contracción con frecuencia es minúscula.
- Tensión-relajación del músculo liso

Otra característica importante del músculo liso, especialmente del tipo unitario visceral de músculo liso de muchos órganos huecos, es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado. Cuando se produce una reducción súbita de volumen, la presión disminuye drásticamente al principio, aunque después aumenta en un plazo de otros pocos segundos o minutos hasta el nivel original o casi hasta el mismo. Estos fenómenos se denominan tensión-relajación y tensión-relajación inversa.
- Regulación de la contracción por los iones calcio
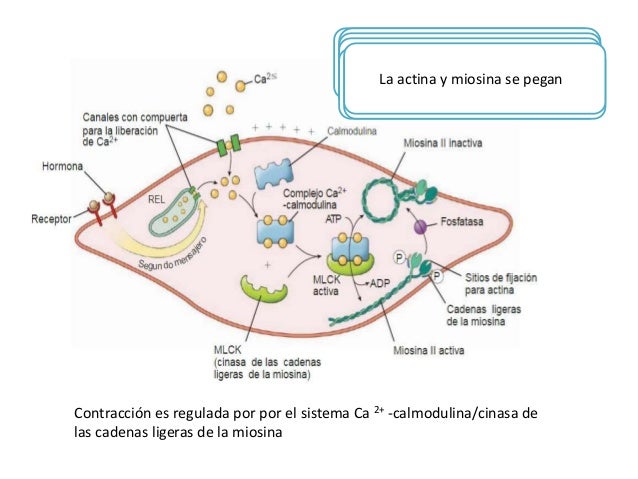
El estímulo que inicia la mayor parte de las contracciones del músculo liso es un aumento de los iones calcio en el medio intracelular. Puede estar producido por la estimulación nerviosa, por estimulación hormonal, por distensión de la fibra o incluso por cambios del ambiente químico de la fibra. El músculo liso no contiene troponina.
Los iones calcio se combinan con la calmodulina para provocar la activación de la miosina cinasa y fosforilación de la cabeza de miosina
En lugar de la troponina, las células musculares lisas contienen una gran cantidad de otra proteína reguladora denominada calmodulina, la cual activa los puentes cruzados de miosina.
La miosina fosfatasa es importante en la interrupción de la contracción
La inversión de la fosforilación de la cabeza de miosina necesita de la enzima miosina fosfatasa, que está localizada en el citosol de la célula muscular lisa y escinde el fosfato de la cadena ligera reguladora. Después se interrumpe el ciclo y finaliza la contracción. El tiempo necesario para la relajación de la contracción muscular está determinado en gran medida por la cantidad de miosina fosfatasa activa en la célula. El número de cabezas unidas a la actina determina la fuerza estática de la contracción.
Control nervioso y hormonal de la contracción del músculo liso
La contracción del músculo liso puede ser estimulada por múltiples tipos de señales:
- Señales nerviosas
- Estimulación hormonal
- Distensión del músculo
- Otros estímulos
La membrana del músculo liso contiene muchos tipos de proteínas receptoras que pueden iniciar el proceso contráctil. Otras proteínas receptoras inhiben la contracción del músculo liso.
Uniones neuromusculares del músculo liso
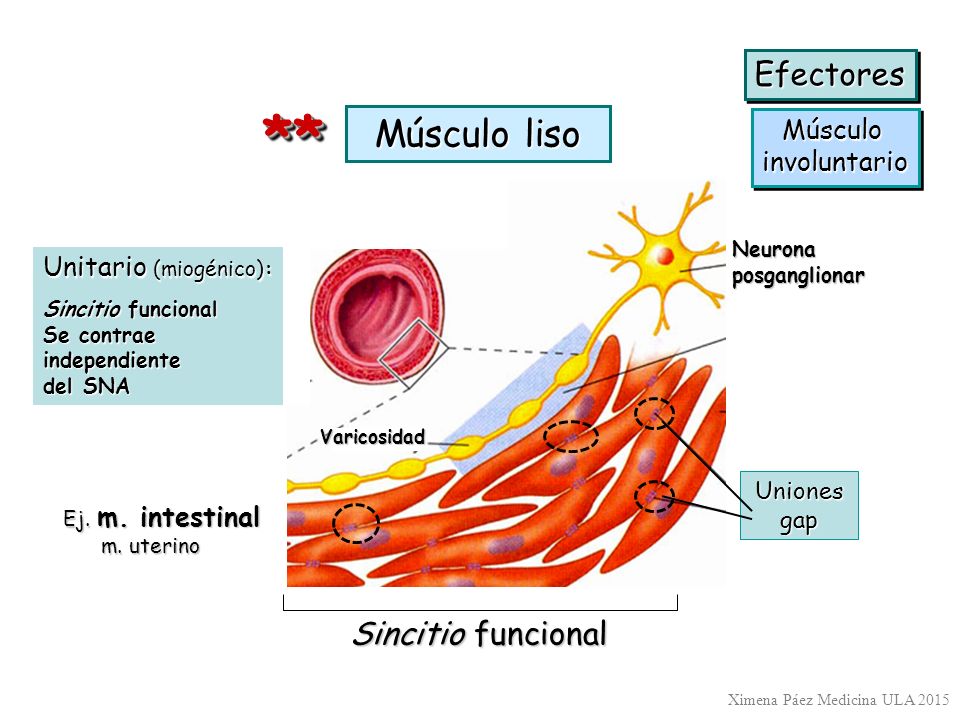
Las ibras nerviosas autónomas que inervan el músculo liso generalmente se ramifican de manera difusa encima de una lámina de fibras musculares. Generalmente no hacen contacto directo con la membrana de las células de las fibras musculares lisas, sino que forman las denominadas uniones difusas que secretan su sustancia transmisora hacia el recubrimiento de matriz del músculo liso. Las fibras nerviosas con frecuencia inervan solo la capa externa, la excitación muscular viaja desde esta capa externa hacia las capas internas por conducción de los potenciales de acción en la masa muscular o mediante difusión adicional de la sustancia transmisora.
Los axones terminales delgados tiene múltiples varicosidades distribuidas a lo largo de sus ejes. En estos puntos se interrumpen las células de Schwann que rodean a los axones, de modo que se puede secretar la sustancia transmisora a través de las paredes de las varicosidades.
Las vesículas de las terminaciones de las fibras nerviosas autónomas contienen acetilcolina en algunas fibras y noradrenalina en otras, y de manera ocasional también otras sustancias. En el tipo multiunitario del músculo liso, las varicosidades están separadas de la membrana de la célula muscular por tan sólo 20 a 30 nm,Estas uniones se denominan uniones de contacto.
Sustancias transmisoras excitadoras e inhibidoras secretadas en la unión neuromuscular del músculo liso
Las sustancias transmisoras más importantes que secretan los nervios autónomos que inervan el músculo liso son acetilcolina y noradrenalina. La acetilcolina es una sustancia transmisora excitadora de las fibras musculares lisas en algunos órganos y un transmisor inhibidor en el músculo liso de otros órganos. Cuando la acetilcolina excita una fibra, la noradrenalina habitualmente la inhibe y de forma inversa.

Tanto a noradrenalina excitan o inhiben el músculo liso uniéndose en primer lugar a una proteína receptora de la superficie de la membrana de la célula muscular. Algunas de las proteínas receptoras son receptores excitadores, mientras que otras son receptores inhibidores.
Potenciales de membrana y potenciales de acción en el músculo liso
- Potenciales de membrana en el músculo liso
El voltaje cuantitativo del potencial de membrana del músculo liso depende de la situación momentánea del músculo. En el estado de reposo normal el potencial intracelular es habitualmente de aproximadamente –50 a –60 mV.
- Potenciales de acción en el músculo liso unitario
Los potenciales de acción del músculo liso visceral se producen por potenciales en espiga y por potenciales de acción con meseta.
Potenciales en espiga
Aparecen en la mayor parte de los tipos de músculo liso unitario. La duración de este tipo de potencial de acción es de 10 a 50 ms. Se dan por estimulación eléctrica, por la acción de hormonas sobre el músculo liso, por la acción de sustancias transmisoras procedentes de las fibras nerviosas, por distensión o como consecuencia de su generación espontánea en la propia fibra muscular.
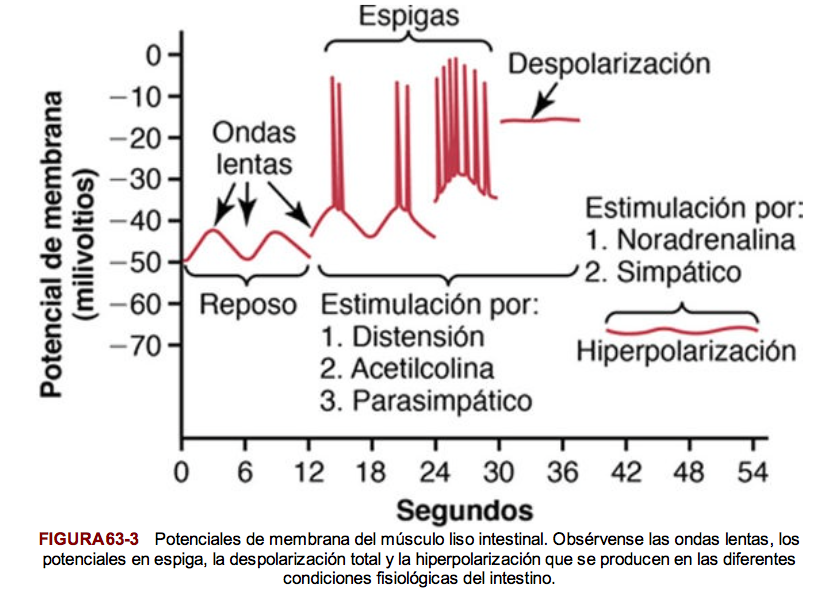
Potenciales de acción con meseta
En lugar de la repolarización rápida de la membrana de la fibra muscular, la repolarización se retrasa durante varios cientos hasta 1.000 ms (1 s). Puede ser responsable de la contracción prolongada que se produce en algunos tipos de músculo liso.

Los canales de calcio son importantes en la generación del potencial de acción del músculo liso
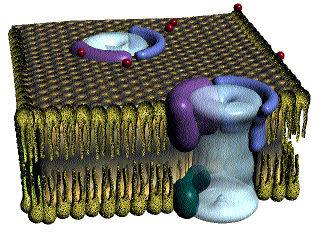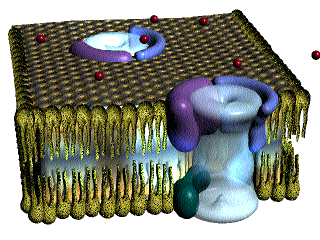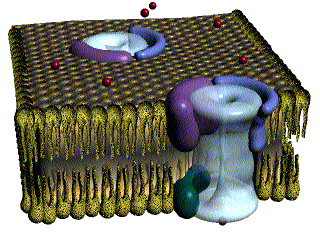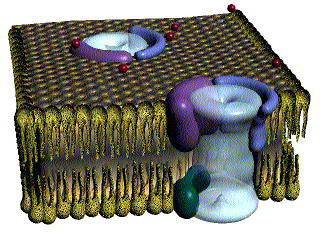
La membrana de la célula muscular lisa tiene muchos más canales de calcio activados por el voltaje, pero pocos canales de sodio. El flujo de iones calcio hacia el interior de la fibra es el principal responsable del potencial de acción. Los canales de calcio se abren muchas veces más lentos que los canales de sodio, y también permanecen abiertos mucho más tiempo.
Los iones calcio actúan directamente sobre el mecanismo contráctil del músculo liso para producir la contracción.
Los potenciales de onda lenta en el músculo liso unitario pueden conducir a la generación espontánea de potenciales de acción
Algunas células musculares lisas son autoexcitadoras. Es decir, los potenciales de acción se originan en las propias células musculares lisas sin ningún estímulo extrínseco. Esto con frecuencia se asocia a un ritmo de ondas lentas básico del potencial de membrana.
Las ondas lentas están producidas por la aparición y desaparición del bombeo de iones positivos hacia el exterior a través de la membrana de la fibra muscular y cuando son lo suficientemente intensas, pueden iniciar potenciales de acción. Cuando el máximo del potencial de la onda lenta negativa en el interior de la membrana celular aumenta en dirección positiva desde –60 hasta aproximadamente –35 mV. Las ondas lentas se denominan ondas marcapasos.
Excitación del músculo liso visceral por distensión muscular
Cuando el músculo liso visceral es distendido lo suficiente, habitualmente se generan potenciales de acción espontáneos, que se deben a una combinación de los potenciales de onda lenta normales y la disminución de la negatividad global del potencial de membrana que produce la propia distensión.
Despolarización del músculo liso multiunitario sin potenciales de acción
Las fibras musculares lisas del músculo liso multiunitario normalmente se contraen sobre todo en respuesta a estímulos nerviosos. Las sustancias transmisoras producen despolarización de la membrana del músculo liso, y esto a su vez produce la contracción.
En las células musculares lisas pequeñas, incluso sin potencial de acción, la despolarización local, denominada potencial de la unión, que produce la propia sustancia transmisora nerviosa se propaga «electrotónicamente» en toda la fibra y es lo único necesario para producir la contracción muscular.
Los efectos de los factores tisulares locales y las hormonas determinan la contracción del músculo liso sin potenciales de acción
- Contracción del músculo liso en respuesta a factores químicos tisulares locales
El músculo liso es muy contráctil y responde rápidamente a los cambios de las condiciones químicas locales del líquido intersticial circundante. Un potente sistema de control de retroalimentación local controla el flujo sanguíneo a la zona tisular local. Algunos de los factores de control específicos son los siguientes:
- La ausencia de oxígeno en los tejidos locales produce relajación del músculo liso y, por tanto, vasodilatación.
- El exceso de anhídrido carbónico produce vasodilatación.
- El aumento de la concentración de iones hidrógeno produce vasodilatación.
La adenosina, el ácido láctico, el aumento de los iones potasio, la disminución de la concentración de los iones calcio y el aumento de la temperatura corporal producen vasodilatación local.
- Efectos de las hormonas sobre la contracción del músculo liso
Las hormonas circulantes en la sangre afectan en cierto grado a la contracción del músculo liso, y algunas tienen efectos profundos. Entre las más importantes se encuentran la noradrenalina, la adrenalina, la acetilcolina, la angiotensina, la endotelina, la vasopresina, la oxitocina, la serotonina y la histamina.
Una hormona produce contracción del músculo liso cuando la membrana de la célula muscular contiene receptores excitadores activados por hormonas para esa hormona. Por el contrario, la hormona produce inhibición si la membrana contiene receptores inhibidores para ella.

- Mecanismos de la excitación o la inhibición del músculo liso por hormonas o por factores tisulares locales
Algunos receptores hormonales de la membrana del músculo liso abren canales iónicos de sodio o de calcio y despolarizan la membrana, al igual que ocurre después de la estimulación nerviosa. Algunas veces la contracción o la inhibición del músculo liso es iniciada por hormonas que no producen directamente ningún cambio en el potencial de membrana. En estos casos la hormona puede activar un receptor de membrana que no abre ningún canal iónico, sino que produce un cambio interno de la fibra muscular.
Para inhibir la contracción se sabe que otros mecanismos activan la enzima adenilato ciclasa o guanilato ciclasa de la membrana celular; las porciones de los receptores que sobresalen hacia el interior de las células están acopladas con estas enzimas, dando lugar a la formación de monofosfato cíclico de adenosina (AMPc) o monofosfato cíclico de guanosina (GMPc), denominados segundos mensajeros.
Origen de los iones calcio que causan la contracción a través de la membrana celular y a partir del retículo sarcoplásmico
Aunque el proceso contráctil del músculo liso es actiado por iones calcio, el origen de estos es diferente. El retículo sarcoplásmico está poco desarrollado en la mayor parte del músculo liso. Por el contrario, la mayoría de los iones calcio que producen la contracción entran en la célula muscular desde el líquido extracelular en el momento del potencial de acción o de otro estímulo.
El tiempo necesario para que se produzca esta difusión es en promedio de entre 200 y 300 ms y se denomina período de latencia antes de que comience la contracción.
Función del retículo sarcoplásmico del músculo liso

Las cavéolas son pequeñas invaginaciones de la membrana celular, serían un análogo rudimentario del sistema de túbulos transversos del músculo esquelético. Se piensa que la transmisión de un potencial de acción hacia las cavéolas excita la liberación de iones calcio desde los túbulos sarcoplásmicos próximos. Mientras más extenso sea el retículo sarcoplásmico de la fibra muscular lisa, más rápidamente se contraerá.
La contracción del músculo liso depende de la concentración extracelular de los iones calcio
Cuando la concentración de iones calcio en el líquido extracelular disminuye a aproximadamente 1/3 a 1/10 de su valor normal, habitualmente se interrumpe la contracción del músculo. La fuerza de la contracción del músculo liso depende mucho de la concentración de iones calcio en el líquido extracelular.
Es necesaria una bomba de calcio para producir la relajación del músculo liso
Para producir la relajación del músculo liso después de que se haya contraído se deben retirar los iones calcio de los líquidos intracelulares. Esta eliminación se consigue mediante una bomba de calcio que bombea iones calcio hacia el exterior de la fibra muscular lisa de nuevo hacia el líquido extracelular o hacia el retículo sarcoplásmico, si está presente.
