
El sistema nervioso: Los Sentidos Especiales.
Capítulo 50: El ojo I. Óptica de la visión
Principios físicos de la óptica
La luz viaja a través de los objetos transparentes a menor velocidad que en e aire. El índice de refracción de una sustancia transparente es el cociente entre la velocidad de la luz en el aire y la velocidad en ese material.
El sentido de la luz siempre es perpendicular al plano del frente de onda.
Aplicación de los principios de la refracción a las lentes
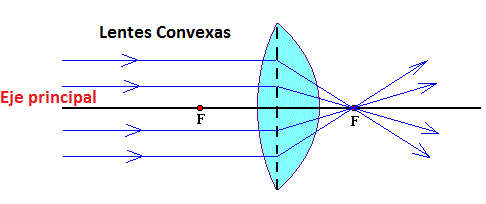
Una lente convexa concentra los rayos de luz.
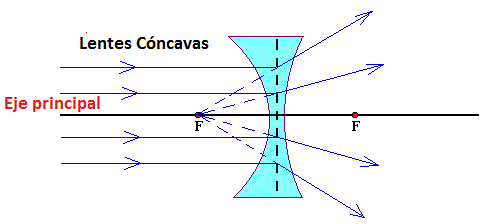
Una lente cóncava dispersa los rayos de luz.
Distancia focal. es la situada más allá de una lente convexa, en la que los rayos de luz paralelos convergen en un punto común. El objeto se ve boca abajo e invertido de izquierda a derecha.

Determinación del poder dióptrico de una lente: «dioptría»
Cuanto más amplia sea la desviación de los rayos luminosos por una lente, mayor es su «poder dióptrico» o poder de refracción. Este poder dióptrico se mide en dioptrías. En el caso de una lente convexa es igual a 1 m dividido por su distancia focal. Por tanto, una lente esférica que cause la convergencia de los rayos luminosos paralelos en un punto focal a 1 m de distancia tiene un poder dióptrico de +1 dioptría. Si la lente es capaz de desviarlos el doble que una lente con un poder de +1 dioptría, se dice que posee una potencia de +2 dioptrías, y los rayos de luz llegan a un punto focal que queda a 0,5 m de la lente. Una lente que provoque la convergencia de los rayos de luz paralelos en un punto focal alejado tan solo 10 cm (0,1m) presenta un poder dióptrico de +10 dioptrías.
Óptica del ojo
El ojo como una cámara
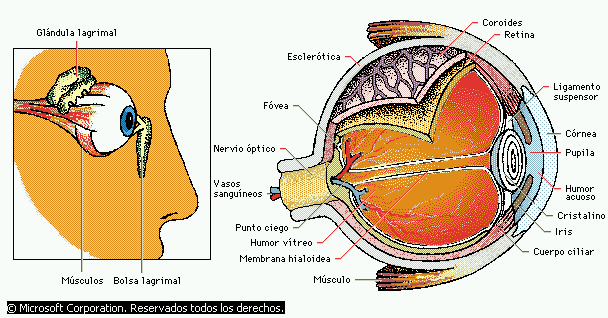
El ojo equivale a una cámara fotográfica corriente desde el punto de vista óptico. Posee un sistema de lentes, un sistema de apertura variable (la pupila) y una retina que corresponde a la película. El sistema ocular de lentes está compuesto por cuatro superficies de refracción:

- La separación entre el aire y la cara anterior de la córnea.
- La separación entre la cara posterior de la córnea y el humor acuoso.
- La separación entre el humor acuoso y la cara anterior del cristalino.
- la separación entre la cara posterior del cristalino y el humor vítreo.
El índice de refracción para el aire es 1; el de la córnea, 1,38; el del humor acuoso, 1,33; el del cristalino (como promedio), 1,4, y el del humor vítreo, 1,34.
Consideración de todas las superficies oculares de refracción como una sola lente: «reducción» del ojo
Si todas las superficies oculares de refracción se suman algebraicamente y a continuación se tratan como una sola lente, la óptica del ojo normal puede simplificarse y representarse de forma esquemática en una «reducción del ojo».
Esta representación resulta práctica para realizar cálculos sencillos. En la reducción del ojo se considera que existe una sola superficie de refracción, con su punto central 17 mm por delante de la retina y un poder dióptrico total de 59 dioptrías cuando la acomodación del cristalino corresponde a la visión de lejos. La cara anterior de la córnea (y no el cristalino) aporta aproximadamente dos tercios de las 59 dioptrías del poder dióptrico ocular.

El índice de refracción de la córnea es sensiblemente distinto al del aire, mientras que el del cristalino no presenta grandes diferencias con los índices del humor acuoso y el humor vítreo.
El poder dióptrico total del cristalino en el interior del ojo, bañado normalmente por líquido a ambos lados, solo es de 20 dioptrías, más o menos la tercera parte del poder dióptrico ocular total. Sin embargo, la importancia de este elemento radica en que, como respuesta a las señales nerviosas procedentes del encéfalo, su curvatura puede aumentar notablemente para permitir la «acomodación».
Formación de una imagen en la retina

De la misma manera que una lente de vidrio es capaz de enfocar una imagen sobre una hoja de papel, el sistema ocular de lentes puede enfocar una imagen sobre la retina. El resultado está dado la vuelta e invertido con respecto al objeto. Sin embargo, la mente percibe los objetos en su posición derecha a pesar de su orientación al revés en la retina debido a que el cerebro está entrenado para considerar como normal una imagen invertida.
Mecanismo de «acomodación»
En los niños, el poder dióptrico del cristalino puede aumentar a voluntad desde 20 dioptrías hasta unas 34, lo que corresponde a una «acomodación» de 14 dioptrías. Para conseguirlo, su forma cambia desde una lente con una convexidad moderada hasta una lente muy convexa. En una persona joven, el cristalino está compuesto por una potente cápsula elástica rellena de un líquido viscoso de carácter proteináceo, pero transparente.

Cuando se encuentra en estado de relajación, sin ninguna tensión aplicada sobre la cápsula, adopta una forma casi esférica, debido básicamente a la retracción elástica de este elemento. Sin embargo, unos 70 ligamentos suspensorios se fijan radialmente en torno al cristalino, y tiran de sus extremos hacia el perímetro exterior del globo ocular. Estos ligamentos se encuentran constantemente tensos por sus inserciones en los bordes anteriores de la coroides y de la retina.
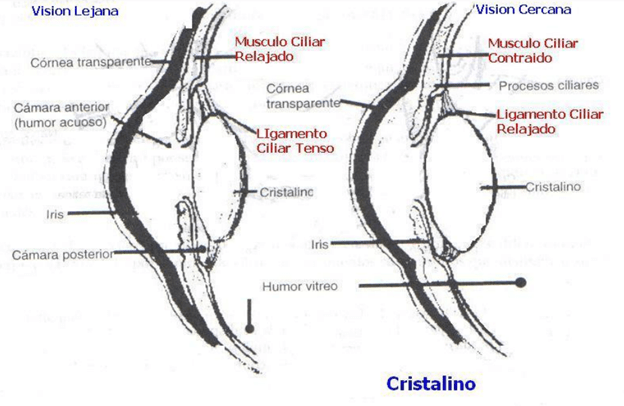
Esta situación hace que el cristalino permanezca relativamente plano si el ojo está en condiciones normales. Además, a nivel de las inserciones laterales de los ligamentos del cristalino en el globo ocular también está situado el músculo ciliar, que posee dos juegos independientes de fibras musculares lisas: las fibras meridionales y las fibras circulares. Las fibras meridionales se extienden desde el extremo periférico de los ligamentos suspensorios hasta la unión esclerocorneal.

Cuando se contraen, arrastran las inserciones periféricas de los ligamentos del cristalino en entido medial hacia los bordes de la córnea, lo que relaja la tensión que ejercen sobre el propio cristalino. Las otras fibras adoptan una disposición circular alrededor de todas las inserciones ligamentosas de modo que, cuando se contraen, producen una acción de tipo esfínter, que reduce el diámetro del perímetro formado por estas inserciones; esta acción también permite que los ligamentos tiren menos de la cápsula del cristalino.

Por tanto, la contracción de cualquiera de los grupos de fibras musculares lisas que forman el músculo ciliar relaja los ligamentos que llegan a la cápsula del cristalino y este último adquiere una forma más esférica, similar a un globo, debido a la elasticidad natural de esta cápsula.
La acomodación está controlada por nervios parasimpáticos
El músculo ciliar está controlado casi en su integridad por señales nerviosas parasimpáticas transmitidas hacia el ojo desde el núcleo del tercer par en el tronco del encéfalo a través de este nervio. La estimulación de los nervios parasimpáticos contrae los dos tipos de fibras que componen el músculo ciliar, lo que relaja los ligamentos del cristalino y propicia un aumento del grosor y del poder dióptrico de dicha estructura. Con el incremento del poder dióptrico, el ojo enfoca objetos más cercanos que cuando posee un poder menor. Por consiguiente, a medida que se aproxima un objeto distante hacia el ojo, la cantidad total de impulsos parasimpáticos que inciden sobre el músculo ciliar ha de crecer de forma progresiva para mantener el objeto constantemente enfocado.
Presbicia: pérdida de acomodación en el cristalino
A medida que una persona envejece, el cristalino crece y se engruesa perdiendo mucha elasticidad, en parte debido a la desnaturalización progresiva de sus proteínas.

La capacidad que posee de modificar su forma disminuye con la edad. El poder de acomodación desciende de unas 14 dioptrías en un niño hasta menos de 2 para la época en que una persona llega a los 45 o 50 años y desciende prácticamente hasta 0 dioptrías con 70 años de edad. A partir de entonces, el cristalino queda casi totalmente desprovisto de su capacidad de acomodación, situación que se conoce como presbicia.
Errores de refracción

Hipermetropía
La hipermetropía suele deberse a la presencia de un globo ocular demasiado corto o, a veces, de un sistema de lentes demasiado débil.
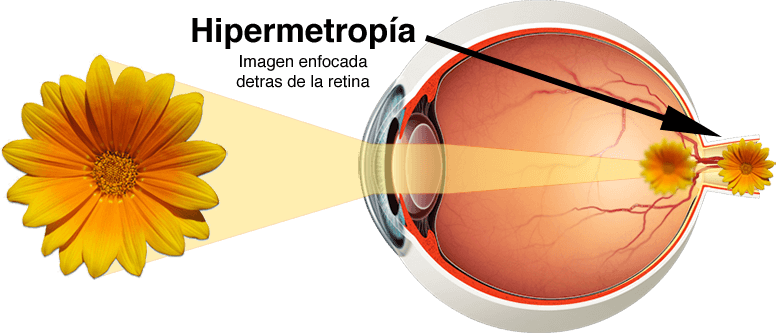
En este proceso el sistema de lentes relajado no desvía lo suficiente los rayos de luz paralelos como para que lleguen a estar enfocados en el momento de alcanzar la retina.
Para vencer esta anomalía, el músculo ciliar ha de contraerse con el fin de incrementar la potencia del cristalino. Una persona hipermétrope, cuando recurre al mecanismo de la acomodación, es capaz de enfocar los objetos alejados en la retina.
Si no ha consumido más que una pequeña parte de la potencia que ofrece el músculo ciliar para acomodar la vista a los objetos distantes, aún le queda un gran poder de acomodación, y también puede enfocar con nitidez otros objetos cada vez más cercanos al ojo hasta llegar al límite de contracción del músculo ciliar.
Miopía
En la miopía, cuando el músculo ciliar está relajado del todo, los rayos de luz procedentes de objetos alejados quedan enfocados delante de la retina.
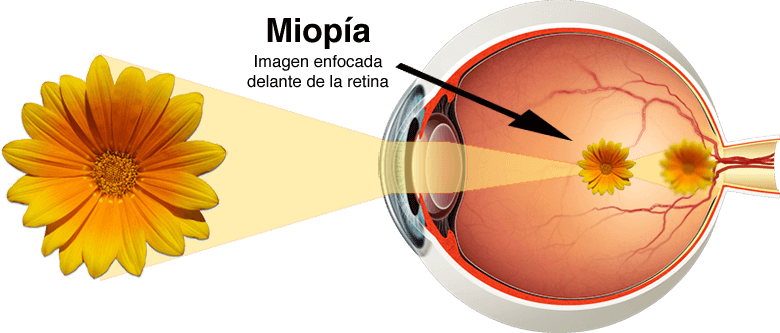
Este trastorno suele deberse a la existencia de un globo ocular demasiado largo, pero puede obedecer a la acción de un poder dióptrico excesivo en el sistema ocular de lentes.
No existe ningún mecanismo por el que el ojo sea capaz de reducir la potencia del cristalino por debajo de la que existe cuando el músculo ciliar se encuentra relajado por completo.
Las personas miopes (o «cortas de vista») carecen de una herramienta para enfocar con nitidez los objetos alejados sobre la retina. Sin embargo, a medida que se acerca el objeto al ojo, acaba por aproximarse lo suficiente como para poder enfocar su imagen. Más tarde, si llega aún más cerca del ojo, puede apelarse al mecanismo de la acomodación para conservar la imagen enfocada con claridad.
Una persona miope posee un «punto lejano» concreto que es el límite para la visión nítida.
Astigmatismo
El astigmatismo es un error de la refracción ocular que hace que la imagen visual de un plano quede enfocada a una distancia diferente de la que corresponde a su plano perpendicular.
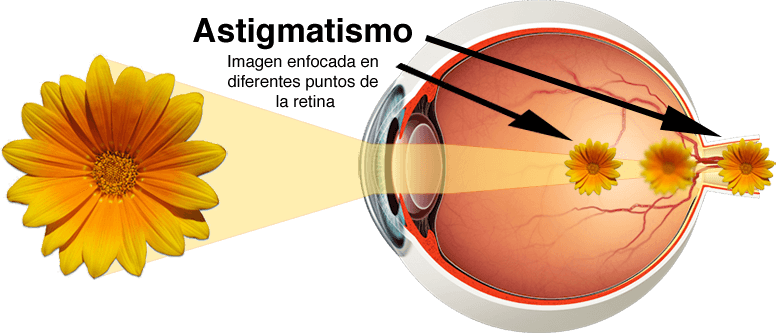
Lo más frecuente es que el astigmatismo obedezca a una curvatura de la córnea demasiado grande en uno de los planos del ojo. Un ejemplo de lente astigmática sería una superficie como la de un huevo situado de lado frente a la luz incidente.
Es evidente que no todos los rayos de luz que atraviesan una lente astigmática llegan a un punto focal común, pues los que cruzan un plano quedan enfocados a una distancia por delante de los que cruzan el otro.
El poder de acomodación del ojo nunca es capaz de compensar el astigmatismo porque su acción cambia la curvatura del cristalino aproximadamente lo mismo en ambos planos; por tanto, en este proceso, cada uno de los dos planos requiere un grado diferente de acomodación. Así pues, sin la ayuda de unas gafas una persona con astigmatismo jamás puede ver las cosas enfocadas con nitidez.
Cataratas: zonas opacas en el cristalino
Las «cataratas» son una alteración ocular especialmente frecuente que sucede sobre todo en las personas mayores. Consiste en una o varias zonas turbias u opacas en el interior del cristalino.
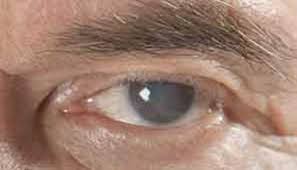
Durante la primera etapa de su formación se desnaturalizan las proteínas de algunas fibras del cristalino. Más adelante, estas mismas proteínas se coagulan para generar áreas opacas en lugar de las fibras proteicas transparentes normales.
Cuando una catarata ha oscurecido la transmisión de luz tanto como para afectar seriamente la visión, la situación puede corregirse mediante la extirpación quirúrgica del cristalino.
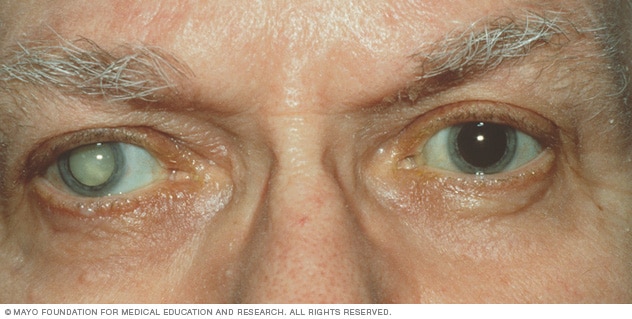
Si se lleva a cabo esta intervención, el ojo pierde una gran parte de su poder dióptrico, que ha de restituirse mediante la colocación de una lente convexa potente delante del mismo; sin embargo, por lo común se implanta una lente artificial de plástico dentro del ojo en el lugar que ocupaba el cristalino extraído.
Agudeza visual
En teoría, la luz procedente de una fuente puntual alejada, cuando se enfoca sobre la retina, debería ser infinitamente pequeña. Sin embargo, dado que el sistema ocular de lentes nunca es perfecto, dicho punto retiniano corrientemente posee un diámetro total de unos 11 μm, incluso cuando el ojo presenta un sistema óptico normal dotado de su máxima resolución. La mancha resulta más brillante en su centro y se va oscureciendo gradualmente hacia los bordes. El diámetro medio de los conos en la fóvea de la retina, que es su porción central, donde la visión está más desarrollada, es de unos 1,5 μm, lo que supone la séptima parte del diámetro del punto luminoso. No obstante, como la mancha de luz tiene un núcleo central brillante y se difumina hacia los bordes, una persona normalmente puede distinguir dos puntos separados si su centro queda a un mínimo de 2 μm de distancia en la retina, lo que excede ligeramente la anchura de los conos en la fóvea. La agudeza visual normal del ojo humano que permite distinguir entre las fuentes puntuales de luz es de unos 25 s de arco. Es decir, cuando los rayos luminosos procedentes de dos puntos distintos chocan con el ojo formando un ángulo mínimo de 25 s entre ellos, normalmente pueden identificarse como dos puntos en vez de uno. Esto significa que una persona con una agudeza visual normal que mire dos minúsculos puntos brillantes de luz a 10 m de distancia apenas puede distinguirlos como entidades independientes cuando estén separados por 1,5 a 2 mm. La fóvea mide menos de 0,5 mm (<500 μm) de diámetro, lo que quiere decir que la agudeza visual máxima ocupa menos de 2° del campo visual. Fuera de esta zona, se va perdiendo agudeza poco a poco, siendo más de 10 veces menor al acercarse a la periferia. Este fenómeno está ocasionado por el número cada vez mayor de conos y bastones que quedan conectados a cada fibra del nervio óptico en las porciones más periféricas de la retina tras abandonar la fóvea.
Sistema humoral del ojo: líquido intraocular
El ojo está relleno de líquido intraocular, que mantiene una presión suficiente en el globo ocular para que siga estando dilatado.

El humor acuoso, que se halla delante del cristalino, y el humor vítreo, que está entre la cara posterior del cristalino y la retina.

El humor acuoso es un líquido que circula con libertad, mientras que el humor vítreo, a veces denominado cuerpo vítreo, es una masa gelatinosa cuya cohesión se mantiene por una fina red fibrilar compuesta básicamente por moléculas de proteoglucanos muy largas. Tanto el agua como las sustancias disueltas pueden difundir con lentitud por el humor vítreo, pero el flujo de líquido es escaso. El humor acuoso se está formando y reabsorbiendo constantemente.

El balance entre su formación y su reabsorción regula el volumen y la presión totales del líquido intraocular.
Formación del humor acuoso por el cuerpo ciliar
El humor acuoso se forma en el ojo a una velocidad media de 2 a 3 ml/min. Básicamente se segrega en su integridad por los procesos ciliares, unos pliegues lineales que sobresalen desde el cuerpo ciliar hacia el espacio que queda detrás del iris donde se fijan los ligamentos del cristalino y el músculo ciliar en el globo ocular.

Debido a su arquitectura plegada, el área total que posee la superficie de los procesos ciliares mide unos 6 cm2 en cada ojo, una gran extensión si se tiene en cuenta el pequeño tamaño del cuerpo ciliar. Las superficies de estos procesos están cubiertas por unas células epiteliales de carácter muy secretor, y justo por debajo de ellas queda una zona muy vascular.
El humor acuoso se forma casi por completo mediante un mecanismo de secreción activa por el epitelio de los procesos ciliares. Este proceso comienza con el transporte activo de los iones sodio hacia los espacios que quedan entre las células. Su paso arrastra con ellos a los iones cloruro y bicarbonato para mantener la neutralidad eléctrica.
A continuación, todos estos iones sumados provocan el desplazamiento osmótico del agua desde los capilares sanguíneos que se hallan debajo de los mismos espacios intercelulares en el epitelio, y la solución resultante fluye desde estos espacios de los procesos ciliares hacia la cámara anterior del ojo. Por ende, diversos nutrientes atraviesan el epitelio por transporte activo o difusión facilitada; entre ellos figuran los aminoácidos, el ácido ascórbico y la glucosa.
Salida del humor acuoso desde el ojo
Una vez que se ha formado el humor acuoso en los procesos ciliares, primero fluye a través de la pupila hacia la cámara anterior del ojo. Desde aquí, circula por delante del cristalino y hacia el ángulo que queda entre la córnea y el iris, después sigue por una trama de trabéculas y finalmente entra en el conducto de Schlemm, que desemboca en las venas extraoculares.

El conducto de Schlemm es una vena de paredes delgadas que recorre el perímetro alrededor del ojo en su integridad. Tiene una membrana endotelial tan porosa que incluso las moléculas grandes de proteínas pueden pasar desde la cámara anterior hacia el conducto de Schlemm, lo mismo que las pequeñas partículas materiales con un tamaño hasta el de los eritrocitos.
Aunque en realidad se trate de un vaso sanguíneo venoso, normalmente es tan grande la cantidad de humor acuoso que entra en el conducto de Schlemm que este líquido lo llena por completo en vez de la sangre. Los pequeños vasos que se extienden desde el conducto de Schlemm hasta las venas más grandes del ojo solo suelen contener humor acuoso, y se denominan venas acuosas.
Presión intraocular
La presión intraocular normal media es de unos 15 mmHg, con un intervalo desde 12 hasta 20 mmHg.
Capítulo 51. El ojo: II. Función receptora y nerviosa de la retina
La retina es la porción del ojo sensible a la luz que contiene:
- Los conos, responsables de la visión de los colores.
- Los bastones, que pueden detectar luz tenue y están encargados básicamente de la visión en blanco y negro y de la visión en la oscuridad.

Ante la excitación de cualquiera de estas células, los impulsos se transmiten primero por la retina a través de las sucesivas capas de neuronas y, finalmente, siguen hacia las fibras del nervio óptico y la corteza cerebral. En este capítulo se explican los mecanismos por los que los conos y los bastones detectan la luz y el color y convierten una imagen visual en las señales del nervio óptico.
Anatomía y función de los elementos estructurales de la retina

Capas de la retina
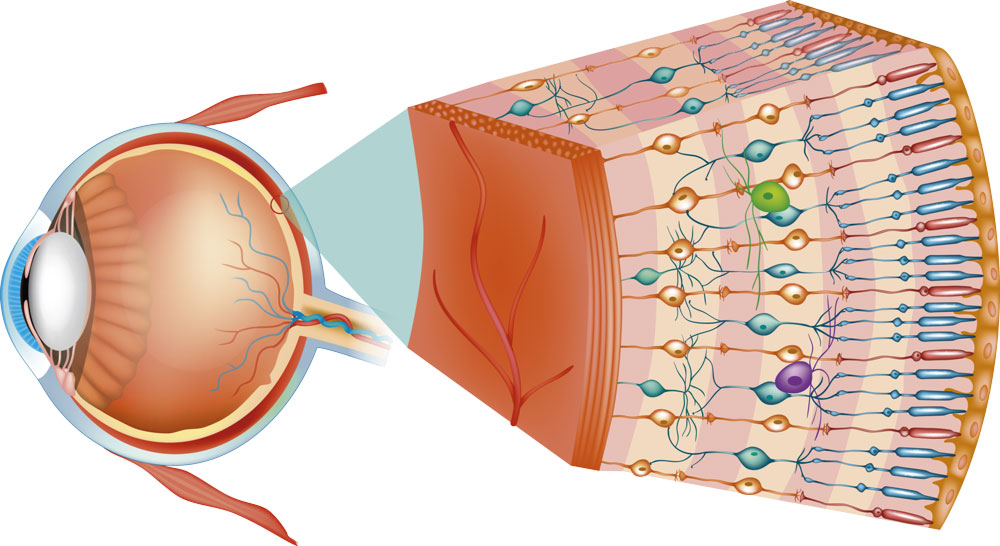
Componentes funcionales de la retina que se disponen en las siguientes capas desde el exterior hacia el interior:
1) capa pigmentaria
2) capa de conos y bastones que aloja las prolongaciones de estos receptores hacia la capa anterior
3) capa nuclear externa que contiene los somas de los conos y los bastones
4) capa plexiforme externa
5) capa nuclear interna
6) capa plexiforme interna
7) capa ganglionar
8) capa de las fibras del nervio óptico
9) membrana limitante interna.

Una vez que la luz atraviesa el sistema ocular de lentes y después el humor vítreo, penetra en la retina desde su interior es decir, pasa primero a través de las células ganglionares y después recorre las capas plexiformes y nucleares antes de llegar finalmente a la capa de los conos y los bastones situada a lo largo de todo el borde externo de la retina. Esta distancia tiene un grosor de varios cientos de micrómetros; la agudeza visual baja debido a este trayecto por un tejido tan poco homogéneo. Sin embargo, según se comenta más adelante, en la región central de la fóvea de la retina las capas internas de la retina quedan apartadas para atenuar esta pérdida de agudeza.
Región de la fóvea de la retina y su importancia en la visión aguda
La fóvea es una zona diminuta en el centro de la retina que ocupa un área total un poco mayor de 1 mm2 se encuentra especialmente capacitada para la visión aguda y detallada.

La fóvea central, de solo 0,3 mm de diámetro, está compuesta por conos casi en su integridad. Estos elementos poseen una estructura especial que contribuye a la detección de detalles en la imagen visual, es decir, los conos de la fóvea presentan un cuerpo especialmente largo y delgado, en contraposición a los conos situados hacia la periferia de la retina, que son mucho más gruesos.
Asimismo, en la región de la fóvea los vasos sanguíneos, las células ganglionares, la capa de células nuclear interna y las capas plexiformes quedan desplazadas hacia un lado en vez de apoyarse directamente sobre la parte superior de los conos, lo que permite que la luz llegue hasta estos sin ningún impedimento.
Conos y bastones
En las porciones periféricas de la retina, los bastones alcanzan un diámetro de 2 a 5 μm, mientras que los conos miden de 5 a 8; en la parte central de la retina, la fóvea, hay bastones, y los conos son delgados y su diámetro solo es de 1,5 μm.

Los principales segmentos funcionales de un cono o de un bastón:
1) el segmento externo.
2) el segmento interno.
3) el núcleo.
4) el cuerpo sináptico.
La sustancia fotosensible se sitúa en el segmento externo.
En el caso de los bastones, esta sustancia fotosensible es la rodopsina; en los conos, es una de las tres sustancias fotosensibles al «color», que suelen denominarse simplemente pigmentos del color, y funcionan casi exactamente igual que la rodopsina excepto por sus diferencias de sensibilidad dentro del espectro.
Cada disco en realidad es un estrato replegado de membrana celular. En cada cono o bastón llega a haber hasta 1.000 discos. Tanto la rodopsina como los pigmentos de color son proteínas conjugadas. Se incorporan a las membranas de los discos bajo la forma de proteínas transmembrana. Las concentraciones de estos pigmentos fotosensibles en los discos son tan grandes que ellos mismos constituyen aproximadamente el 40% de toda la masa del segmento externo.
El segmento interno del cono o del bastón contiene el citoplasma habitual con los orgánulos citoplásmicos. Especialmente importantes son las mitocondrias que, según se explica más adelante, cumplen la decisiva misión de proporcionar energía para el funcionamiento de los fotorreceptores. El cuerpo sináptico es la porción del bastón o del cono que conecta con las siguientes estructuras neuronales, las células horizontales y bipolares, que representan las siguientes etapas en la cadena visual.
Capa pigmentaria de la retina
El pigmento negro melanina de la capa pigmentaria impide la reflexión lumínica por toda la esfera del globo ocular, lo que resulta importantísimo para una visión nítida. El pigmento cumple la misma función en el ojo que el color negro en el interior de los fuelles de una cámara. Sin él, los rayos de luz se reflejarían en todas las direcciones dentro del globo y generarían una iluminación difusa de la retina en vez del contraste normal entre los puntos claros y oscuros necesario para la formación de una imagen precisa.

La trascendencia de la melanina en la capa pigmentaria queda bien patente por su ausencia en los albinos (personas que carecen de ella en cualquier zona de su cuerpo por causas hereditarias). Cuando un albino entra en una habitación radiante, la luz que incide sobre la retina se refleja en todas las direcciones dentro del globo ocular por sus superficies internas desprovistas de pigmentos y por la esclerótica subyacente, de modo que un solo punto de luz aislado que en condiciones normales no excitaría nada más que a unos pocos conos y bastones, se refleja por todas partes y activa muchos receptores.

Por tanto, la agudeza visual de los albinos, incluso con la mejor corrección óptica, rara vez supera los 20/100 a 20/200 en vez de los valores normales de 20/20. La capa pigmentaria también alberga grandes cantidades de vitamina A. Esta sustancia se intercambia hacia dentro y hacia fuera a través de las membranas celulares en los segmentos externos de los conos y los bastones, que están a su vez inmersos en el pigmento.
Fotoquímica de la visión
Tanto los conos como los bastones contienen productos químicos que se descomponen ante la exposición a la luz y, en el curso del proceso, excitan a las fibras nerviosas que salen del ojo. La sustancia sensible a la luz en los bastones se llama rodopsina; en los conos, donde se denominan pigmentos de los conos o pigmentos del color, su composición no es nada más que un poco diferente a la que presenta la rodopsina.
Ciclo visual rodopsina-retinal y excitación de los bastones
La rodopsina y su descomposición por la energía lumínica
El segmento externo de los bastones que se extiende hacia la capa pigmentaria de la retina presenta una concentración aproximada del 40% del pigmento sensible a la luz llamado rodopsina, o púrpura visual. Esta sustancia es una combinación de la proteína escotopsina y el pigmento carotenoide retinal (también llamado «retineno»). Además, el retinal es de un tipo especial denominado 11-cis-retinal. Esta forma cis resulta importante porque solo ella puede unirse a la escotopsina para sintetizar rodopsina.

Cuando la rodopsina absorbe la energía lumínica, empieza a descomponerse en una fracción muy pequeña de segundo.
El origen de esta rápida descomposición se sitúa en la fotoactivación de los electrones situados en la porción retinal de esta sustancia, que desemboca en la transformación instantánea de la forma cis del retinal en una forma todo-trans que conserva la misma estructura química que la forma cis pero presenta una estructura física diferente: es una molécula recta en vez de plegada.
Como la orientación tridimensional de los lugares reactivos en el todo-trans-retinal deja de coincidir con la orientación correspondiente en la proteína escotopsina, el todo-trans-retinal empieza a soltarse de ella. El producto inmediato es la batorrodopsina, que representa una combinación parcialmente disociada del todo-trans- retinal y la escotopsina. La batorrodopsina resulta sumamente inestable y se degrada en cuestión de nanosegundos en lumirrodopsina.
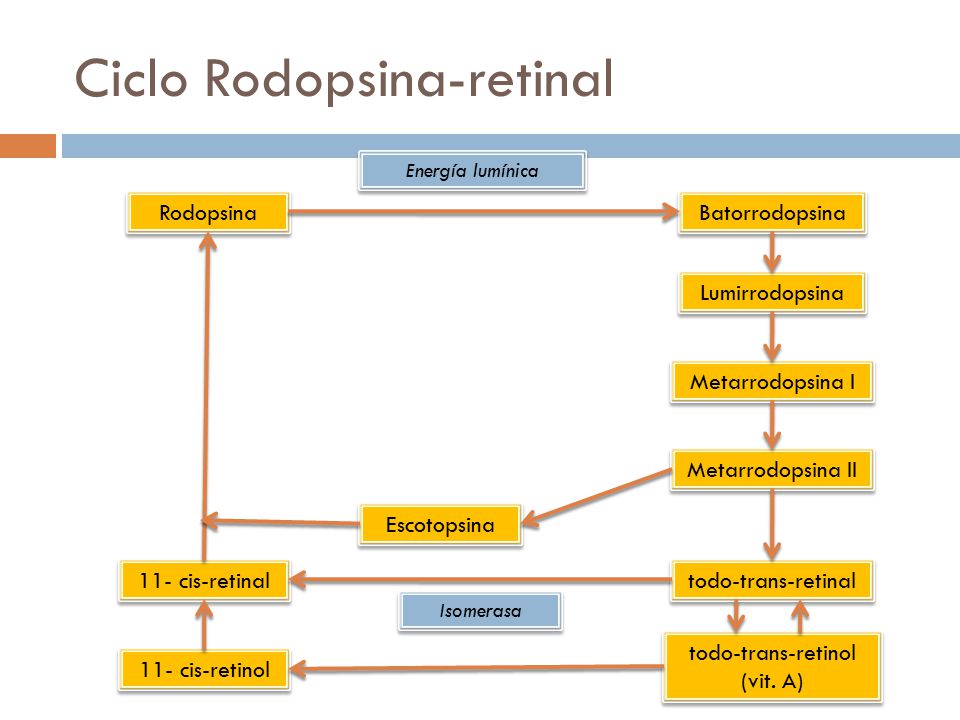
Después de unos microsegundos, este producto se descompone en metarrodopsina I, en 1ms más proximadamente pasa a metarrodopsina II, y por último, con mucha mayor lentitud (en un plazo de segundos), se convierte en los productos escindidos por completo de la escotopsina y el todo-trans-retinal. Es la metarrodopsina II, también llamada rodopsina activada, la que estimula los cambios eléctricos en los bastones, y tras ello estos receptores ya transmiten la imagen visual hacia el sistema nervioso central bajo la forma de un potencial de acción en el nervio óptico.
Regeneración de la rodopsina
La primera etapa en la regeneración de la rodopsina consiste en reconvertir el todo-trans-retinal en 11-cis- retinal. Este proceso requiere energía metabólica y está catalizado por la enzima isomerasa de retinal. Una vez que se ha formado el 11-cis-retinal, automáticamente se recombina con la escotopsina para reconstituir la rodopsina, que a continuación permanece estable hasta que se desencadena una vez más su descomposición por la nueva absorción de energía lumínica.
Excitación del bastón cuando la luz activa la rodopsina
El potencial de receptor del bastón es hiperpolarizante, no despolarizante
Cuando el bastón se encuentra expuesto a la luz, el potencial de receptor resultante es diferente del que existe en casi todos los demás receptores sensitivos, dado que la excitación de un bastón provocar un aumento de la negatividad en el potencial de membrana en su interior, lo que supone un estado de hiperpolarización. Este fenómeno es exactamente lo contrario del descenso de la negatividad (el proceso de «despolarización») que sucede en casi todos los demás receptores sensitivos.
El segmento interno bombea sodio sin interrupción desde el interior del bastón hacia su exterior y se bombean iones potasio hacia el interior de la célula. Los iones potasio se filtran de la célula a través de canales de potasio no activados que están confinados en el segmento interno del bastón. Como en otras células, esta bomba de sodio-potasio crea un potencial negativo dentro de la célula tomada en su integridad. Sin embargo, el segmento externo, donde están situados los discos fotorreceptores, es totalmente diferente; aquí, la membrana del bastón, en situación de oscuridad, resulta permeable para los iones sodio que fluyen a través de canales activados por monofosfato de guanosina cíclico (GMPc). En el estado de oscuridad, los niveles de GMPc son elevados, lo que hace posible que estos iones de carga positiva se filtren permanentemente hacia el interior del bastón y neutralicen así gran parte de la negatividad que hay dentro de toda la célula.

Por tanto, en condiciones de oscuridad normales, cuando el bastón no está excitado, hay una baja electronegatividad en la parte interna a su membrana, que mide unos –40mV en vez de los habituales –70 a –80 presentes en la mayoría de los receptores sensitivos. Cuando la rodopsina del segmento externo del bastón queda expuesta a la luz, se activa y empieza a descomponerse. A continuación, los canales de sodio activados por GMPc se cierran y la conductancia al sodio de la membrana del segmento externo hacia el interior del bastón se reduce a través de un proceso en tres etapas: 1) la luz es absorbida por la rodopsina, lo que provoca la fotoactivación de los electrones en la parte retiniana, según se ha descrito anteriormente; 2) la rodopsina activada estimula una proteína G denominada transducina, que después activa la GMPc fosfodiesterasa, una enzima que cataliza la descomposición de GMPc en 5’-GMPc, y 3) la reducción en GMPc cierra los canales de sodio activados por GMPc y reduce la corriente de sodio hacia el interior. Los iones sodio siguen siendo bombeados hacia fuera a través de la membrana del segmento interno. Por tanto, ahora los iones sodio que salen del bastón son más que los que vuelven a entrar. Como se trata de iones positivos, su pérdida desde el interior crea más negatividad por dentro de la membrana, y cuanto mayor sea la magnitud de la energía lumínica que llega al bastón, más acusada se vuelve la electronegatividad (es decir, mayor es el grado de hiperpolarización).
Con una intensidad máxima de luz, el potencial de membrana se aproxima a –70 o –80 mV, que está cerca del potencial de equilibrio para los iones potasio a través de la membrana.
Duración del potencial de receptor y relación logarítmica entre dicho potencial y la intensidad de la luz
Cuando un pulso repentino de luz alcanza la retina, la hiperpolarización transitoria (potencial de receptor) que se produce en los bastones llega a su máximo en cuestión de unos 0,3s y dura más de 1s. En los conos, el cambio sucede cuatro veces más rápido que en los bastones. Una imagen visual que incida sobre los bastones de la retina tan solo durante una millonésima de segundo a veces puede causar la sensación de verla por un tiempo superior a 1s. Otra característica del potencial de receptor es que resulta aproximadamente proporcional al logaritmo de la intensidad de la luz. Esta característica es importantísima porque le permite al ojo distinguir esta variable a lo largo de un intervalo muchos miles de veces mayor de lo que sería posible en caso contrario.
Mecanismo por el que la descomposición de la rodopsina disminuye la conductancia al sodio de la membrana: la «cascada» de excitación
En condiciones óptimas, un solo fotón de luz, la unidad cuántica de energía lumínica más pequeña posible, puede generar un potencial de receptor de 1 mV aproximadamente en un bastón. Ya con 30 fotones de luz se logrará producir la mitad de la saturación del bastón. Los fotorreceptores poseen una cascada química sumamente sensible que amplifica los efectos estimuladores en torno a un millón de veces, del modo siguiente:
1. El fotón activa un electrón en la porción 11-cis-retinal de la rodopsina; esta activación propicia la generación de metarrodopsina II, que es la forma activa de la rodopsina.
2. La rodopsina activada funciona como una enzima que estimula muchas moléculas de transducina, una proteína presente en su forma inactiva en las membranas de los discos y en la membrana celular de bastón.
3. La transducina activada estimula otras muchas moléculas más de fosfodiesterasa.
4. La fosfodiesterasa activada es otra enzima; su acción hidroliza de inmediato muchas moléculas de GMPc, destruyéndolas.
5. En cuestión de 1s más o menos, otra enzima, la cinasa de rodopsina, que está presente siempre en el bastón, desactiva la rodopsina activada (la metarrodopsina II), y toda la cascada se invierte hasta volver a su estado normal con los canales de sodio abiertos. Por tanto, los bastones han adquirido una importante cascada química que amplifica el efecto de un solo fotón de luz hasta provocar el movimiento de millones de iones sodio. Este mecanismo explica la sensibilidad extrema de estos receptores en condiciones de oscuridad. Los conos son de 30 a 300 veces menos sensibles que los bastones, pero incluso este grado de sensibilidad permite la visión de los colores a cualquier intensidad de luz superior a la de un crepúsculo sumamente tenebroso.
Fotoquímica de la visión de los colores por los conos

Anteriormente se ha señalado que las sustancias fotosensibles de los conos poseen casi exactamente la misma composición química que la rodopsina de los bastones. La única diferencia radica en que sus porciones proteicas u opsinas (llamadas fotopsinas en los conos) son un poco distintas de la escotopsina de los bastones. La porción de todos los pigmentos visuales correspondiente al retinal es absolutamente idéntica en los conos y en los bastones. Por tanto, los pigmentos sensibles al color de los conos consisten en combinaciones de retinal y fotopsinas. En la explicación sobre la visión de los colores que se ofrece más adelante en este capítulo, quedará patente que solo uno de los tres tipos de pigmentos destinados a este fin está presente en cualquiera de los diferentes conos, lo que les concede una sensibilidad selectiva a los diversos colores: azul, verde o rojo. Estos pigmentos para el color se llaman, respectivamente, pigmento sensible al azul, pigmento sensible al verde y pigmento sensible al rojo. Sus características de absorción en los tres tipos de conos muestran unas absorbencias máximas ante la luz de las longitudes de onda de 445, 535 y 570 nm, respectivamente. Estas son también las longitudes de onda con una sensibilidad extrema a la luz en cada tipo de cono, lo que empieza a sentar las condiciones para poder explicar cómo distingue los colores la retina. También se ofrece la curva de absorción de la rodopsina en los bastones, con un pico a 505 nm.
Visión en color
Mecanismo tricolor para la detección del color
Cualquier teoría sobre la visión de los colores está basada en la célebre observación de que el ojo humano es capaz de detectar casi todas las gradaciones de color cuando solo las luces roja, verde y azul monocromáticas se mezclan adecuadamente en diversas combinaciones.

Sensibilidades espectrales de los tres tipos de conos
A partir de las exploraciones sobre visión de los colores, se ha comprobado que las sensibilidades espectrales de los tres tipos de conos son básicamente idénticas en el hombre a las curvas de absorción de la luz para los tres tipos de pigmentos presentes en estas células.
Interpretación del color en el sistema nervioso

Una luz monocromática naranja dotada de una longitud de onda de 580 nm estimula los conos rojos hasta un valor más o menos de 99 (el 99% de la estimulación máxima con la longitud de onda óptima); en el caso de los conos verdes los activa con un valor del estímulo de 42 aproximadamente, pero no causa ningún efecto en absoluto sobre los conos azules. Por tanto, las proporciones de estimulación de los tres tipos de conos en este caso son 99:42:0. El sistema nervioso interpreta esta colección de proporciones como la sensación correspondiente al naranja.

En cambio, una luz monocromática azul con una longitud de onda de 450 nm estimula los conos rojos hasta un valor de 0, los verdes hasta un valor de 0 y los azules hasta un valor de 97. El sistema nervioso interpreta este juego de proporciones (0:0:97) como azul. En el mismo sentido, los cocientes 83:83:0 se interpretan como amarillo y 31:67:36 como verde.
Percepción de la luz blanca
Una estimulación aproximadamente equivalente de los conos rojo, verde y azul da lugar a la sensación visual del blanco.
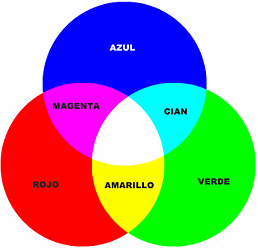
Con todo, la luz no tiene una sola longitud de onda que corresponda al blanco; en su lugar, este color es una combinación de todas las longitudes del espectro. Además, la percepción del blanco puede lograrse mediante la estimulación de la retina con la combinación oportuna tan solo de tres colores escogidos que activen los tipos de conos respectivos más o menos por igual.
Función nerviosa de la retina
Los diversos tipos neuronales son los siguientes:
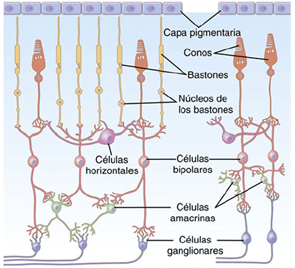
1. Los fotorreceptores, los conos y los bastones, que transmiten las señales hacia la capa plexiforme externa, donde hacen sinapsis con las células bipolares y horizontales.
2. Las células horizontales, que transmiten las señales en sentido horizontal por la capa plexiforme externa desde los conos y los bastones hasta las células bipolares.
3. Las células bipolares, que transmiten las señales en sentido vertical desde los conos, los bastones y las células horizontales hacia la capa plexiforme interna, donde hacen sinapsis con las células ganglionares y amacrinas.
4. Las células amacrinas, que transmiten las señales en dos direcciones, directamente desde las células bipolares hasta las células ganglionares, u horizontalmente en el seno de la capa plexiforme interna desde los axones de las células bipolares hasta las dendritas de las células ganglionares o hasta otras células amacrinas.
5. Las células ganglionares, que transmiten las señales de salida desde la retina hacia el cerebro a través del nervio óptico.
Un sexto tipo neuronal en la retina, que no es muy abundante y está ausente en la figura, es la célula interplexiforme. Este elemento transmite señales en sentido retrógrado desde la capa plexiforme interna hacia la capa plexiforme externa. Su carácter es inhibidor y se cree que controla la dispersión lateral de los impulsos visuales por las células horizontales en esta última capa. Su cometido puede consistir en contribuir a regular el grado de contraste de la imagen visual.
Neurotransmisores liberados por las neuronas de la retina
No se han aclarado por completo todas las sustancias químicas neurotransmisoras que participan en la transmisión sináptica de la retina. Sin embargo, los conos y los bastones liberan glutamato en sus sinapsis con las células bipolares. Los estudios histológicos y farmacológicos han demostrado la existencia de muchas clases de células amacrinas que segregan un mínimo de ocho tipos de sustancias transmisoras, como ácido γ-aminobutírico (GABA), glicina, dopamina, acetilcolina e indolamina, cuya acción normalmente posee en general un carácter inhibidor. Los de las células bipolares, horizontales e interplexiformes no están nada claros, pero al menos parte de las células horizontales liberan transmisores inhibidores.
Células amacrinas y sus funciones
Se han identificado unas 30 clases de células amacrinas por medios morfológicos e histoquímicos.

• Un tipo de célula amacrina forma parte de la vía directa para la visión de los bastones, a saber, la compuesta por el bastón, las células bipolares, las células amacrinas y las células ganglionares.
• Otro tipo de célula amacrina responde potentemente cuando comienza una señal visual continua, pero su actividad se extingue con rapidez.
• Otras células amacrinas presentan una respuesta enérgica al desaparecer las señales visuales, pero, una vez más, su descarga cesa a gran velocidad.
• Otro tipo responde cuando se enciende o se apaga una luz, para indicar simplemente un cambio de iluminación, sin tener en cuenta su sentido.
• Otro tipo más responde al movimiento de un punto a través de la retina en una dirección específica; por tanto, se dice que estas células amacrinas son sensibles a la dirección.
Así pues, y en un cierto sentido, muchas de las células amacrinas o su mayoría son interneuronas que sirven para analizar las señales visuales antes de que lleguen a abandonar la retina.
Células ganglionares y fibras del nervio óptico
Cada retina contiene unos 100 millones de bastones y 3 millones de conos; con todo, el número de células ganglionares solo llega a 1,6 millones más o menos. Por tanto, como promedio son 60 bastones y 2 conos los que convergen sobre cada célula ganglionar y la fibra del nervio óptico que sale desde ella hacia el cerebro. Sin embargo, existen grandes diferencias entre la retina periférica y la retina central. A medida que nos acercamos hacia la fóvea, disminuye la cantidad de conos y de bastones que convergen sobre cada fibra óptica, y los propios receptores también se vuelven más escasos. Estos efectos acentúan progresivamente la agudeza visual en la retina central. En su mismo centro, la fóvea central, no hay más que unos pocos conos finos (en torno a 35.000) y ningún bastón.
Asimismo, el número de fibras del nervio óptico que nace en esta porción de la retina coincide casi exactamente con el número de conos. Este fenómeno explica el enorme grado de agudeza visual que presenta la retina central en comparación con su nivel mucho más limitado en la periferia.
Otra diferencia existente entre las porciones central y periférica de la retina consiste en la sensibilidad mucho mayor de esta última a la luz tenue, lo que obedece en parte al hecho de que los bastones son de 30 a 300 veces más sensibles que los conos. Sin embargo, esta mayor sensibilidad se exagera más si se tiene en cuenta que hasta 200 bastones convergen sobre una sola fibra del nervio óptico en las porciones más periféricas de la retina, de modo que las señales procedentes de estos receptores se suman entre sí para propiciar una estimulación aún más intensa de las células ganglionares periféricas y sus fibras correspondientes en el nervio óptico.
Capitulo 53: El ojo III :Neurofisiología central de la visión.
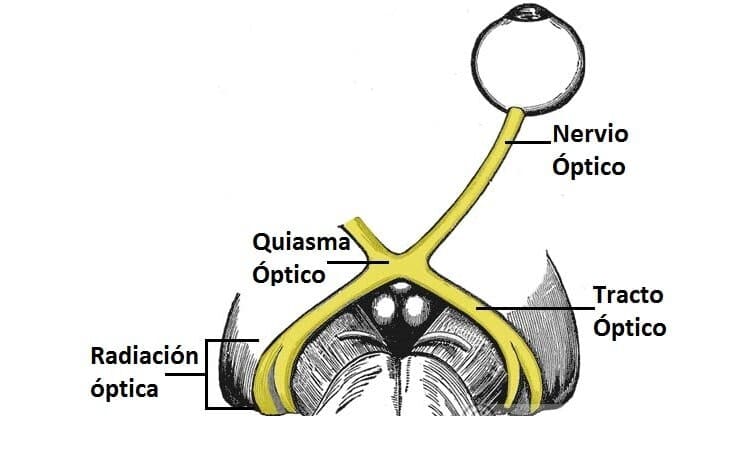
Vías visuales: Las principales vías visuales que van desde las dos retinas hasta la corteza visual. Las señales nerviosas de este carácter abandonan la retina a través de los nervios ópticos.

En el quiasma óptico, las fibras procedentes de la mitad nasal de la retina cruzan hacia el lado opuesto, donde se unen a las fibras originadas en la retina temporal contraria para formar los tractos ópticos o cintillas ópticas. A continuación, las fibras de cada tracto óptico hacen sinapsis en el núcleo geniculado lateral dorsal del tálamo, y desde allí, las fibras geniculocalcarinas se dirigen a través de la radiación óptica (también denominada tracto geniculocalcarino) hacia la corteza visual primaria en el área correspondiente a la cisura calcarina del lóbulo occipital medial.

Las fibras visuales también se dirigen a otras regiones más antiguas del encéfalo:
1) desde los tractos ópticos llega hasta el núcleo supraquiasmático del hipotálamo, se supone que para regular los ritmos circadianos que sincronizan los diversos cambios fisiológicos del organismo según la noche y el día.
2) hacia los núcleos pretectales en el mesencéfalo, para suscitar movimientos reflejos de los ojos a fin de enfocarlos sobre los objetos de importancia y activar el reflejo fotomotor pupilar.
3) hacia el colículo superior, para controlar los movimientos direccionales rápidos de ambos ojos.
4) hacia el núcleo geniculado lateral ventral del tálamo y las regiones basales adyacentes del cerebro, se cree que para contribuir al dominio de algunas de las funciones conductuales que lleva a cabo el organismo.
Anomalías del campo visual.
A veces, los puntos ciegos se encuentran en porciones del campo visual diferentes al área del disco óptico. Tales puntos, llamados escotomas, a menudo están causados por el daño del nervio óptico como consecuencia de un glaucoma (presión excesiva del líquido contenido en el interior del globo ocular), las reacciones alérgicas a nivel de la retina o procesos tóxicos, como el saturnismo o el consumo excesivo de tabaco.

Otro trastorno que puede diagnosticarse con la campimetría es la retinitis pigmentaria. En esta enfermedad, ciertas porciones de la retina degeneran, y en las zonas degeneradas se deposita un exceso del pigmento melanina. La retinitis pigmentaria primero suele ocasionar una ceguera en el campo visual periférico y a continuación invadir gradualmente las áreas centrales.
Campos visuales; campimetría.
El campo visual es la zona de visión observada por un ojo en un instante dado. La región percibida por el lado nasal se llama campo visual nasal y la que llega al lado lateral campo visual temporal.

Para diagnosticar una ceguera en una porción específica de la retina, se cartografía el campo visual de cada ojo mediante un procedimiento llamado campimetría. Esta cartografía se realiza haciendo que el individuo mire con un ojo hacia un punto central situado directamente delante de él; el otro ojo está cerrado.
Efecto de las lesiones de la vía óptica sobre los campos visuales.

La destrucción de todo el nervio óptico origina una ceguera en el ojo afectado. Si el trastorno afecta al quiasma óptico impide el cruce de los impulsos que proceden de la mitad nasal de cada retina y van dirigidos hacia el tracto óptico del lado opuesto. Por tanto, esta mitad queda ciega a ambos lados, lo que significa que la persona está ciega en el campo temporal de cada ojo porque la imagen del campo visual se encuentra invertida en la retina debido al sistema óptico ocular; esta situación se denomina hemianopsia bitemporal. Tales lesiones normalmente son el resultado de un tumor hipofisario que presiona sobre la parte inferior del quiasma desde la silla turca hacia arriba.
Control muscular de los movimientos oculares.

Los movimientos oculares están controlados por tres pares de músculos:
1) los rectos medial y lateral.
2) los rectos superior e inferior.
3) los oblicuos superior e inferior.
Los rectos medial y lateral se contraen para desplazar los ojos de un lado a otro. Los rectos superior e inferior lo hacen para moverlos hacia arriba y hacia abajo. En cuanto a los músculos oblicuos, intervienen sobre todo en la rotación de los globos oculares a fin de mantener los campos visuales en posición vertical.
Control del diámetro pupilar.

La estimulación de los nervios parasimpáticos también excita el músculo esfínter de la pupila, lo que disminuye por esta vía la apertura pupilar; este proceso se denomina miosis. A la inversa, la estimulación de los nervios simpáticos excita las fibras radiales del iris y provoca la dilatación pupilar, lo que se denomina midriasis.
Reflejo pupilar fotomotor.

Cuando la luz ilumina los ojos, las pupilas se contraen, reacción llamada reflejo pupilar fotomotor. La función del reflejo fotomotor consiste en ayudar al ojo a adaptarse de forma rapidísima a unas condiciones lumínicas cambiantes.
El diámetro pupilar tiene unos límites en torno a 1,5 mm por su extremo inferior y a 8 mm por el superior. Por tanto, dado que el brillo de la luz que llega a la retina crece con el cuadrado de esta variable, la amplitud de la adaptación a la luz y a la oscuridad que puede alcanzarse mediante el reflejo pupilar más o menos es de 30 a 1; es decir, la cantidad de luz que penetra en el ojo cambia hasta 30 veces.
Síndrome de Horner.
Los nervios simpáticos del ojo a veces quedan interrumpidos. Esta circunstancia suele suceder en la cadena simpática cervical, lo que provoca el cuadro clínico llamado síndrome de Horner.
Este síndrome ocasiona los siguientes efectos:
1) Debido a la interrupción de las fibras nerviosas simpáticas dirigidas al músculo dilatador de la pupila, esta permanece contraída de forma continua con un diámetro más pequeño que la pupila del lado opuesto.
2) El párpado superior se cae debido a que normalmente se mantiene en posición abierta durante las horas de vigilia en parte por la contracción de las fibras musculares lisas contenidas en su interior e inervadas por el sistema simpático.
Por tanto, la destrucción de estos nervios imposibilita su apertura hasta una altura normal. Tercero, los vasos sanguíneos del lado correspondiente de la cara y de la cabeza quedan dilatados de un modo persistente. Cuarto, no puede producirse la sudoración (que requiere la acción de las señales nerviosas simpáticas) en el mismo lado de la cara y de la cabeza afectado por el síndrome de Horner.
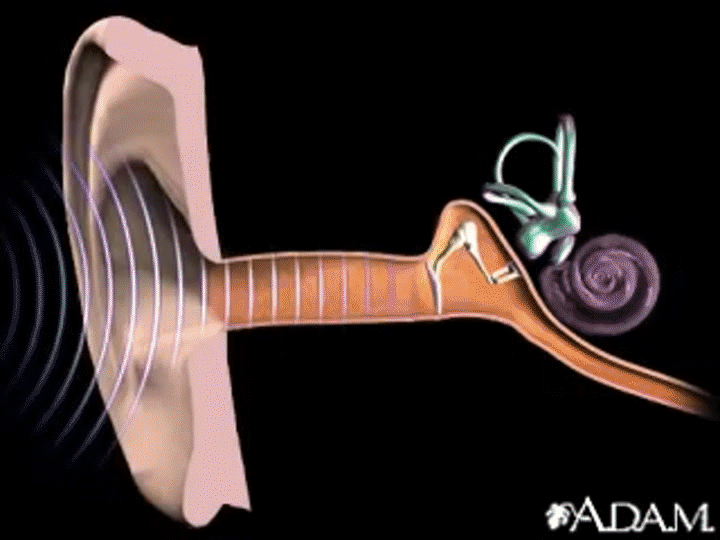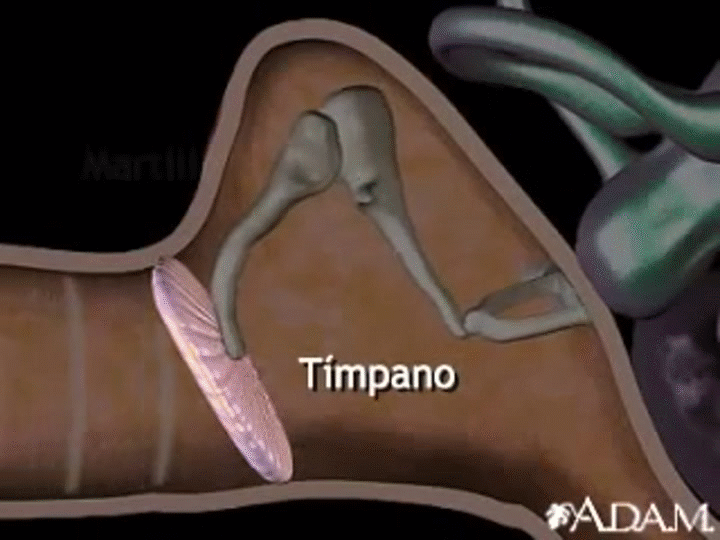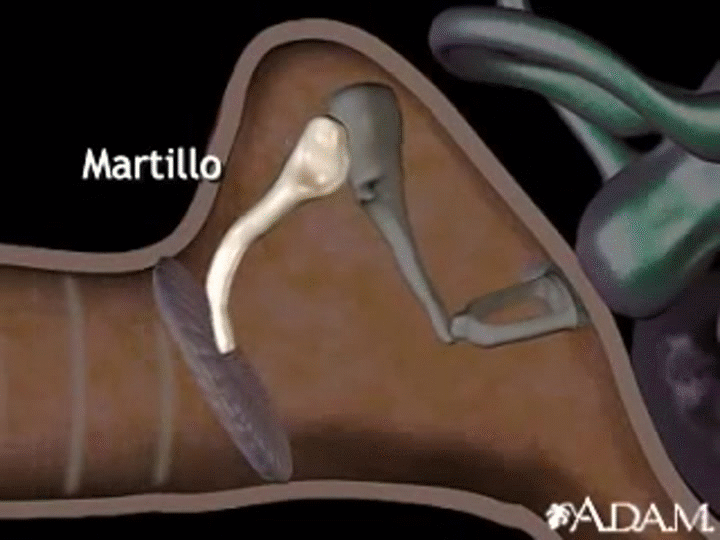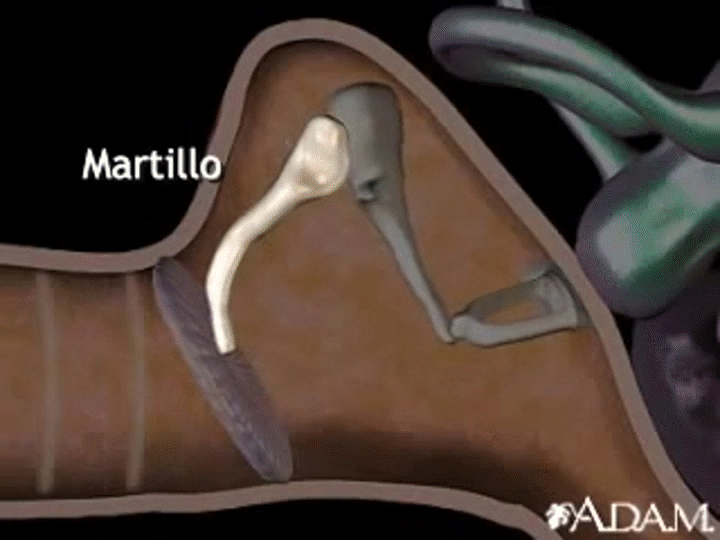
Capítulo 53 : El sentido de la audición
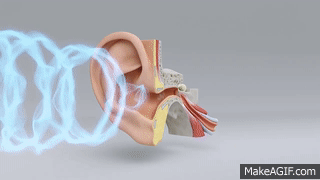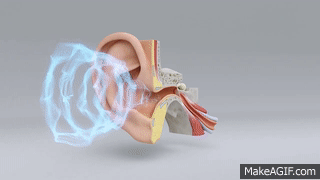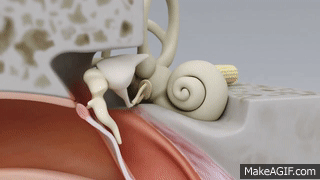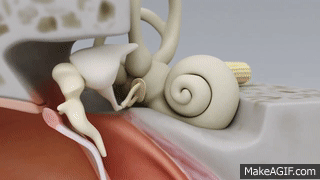
La membrana timpánica y el sistema de huesecillos
La membrana timpánica tiene forma cónica
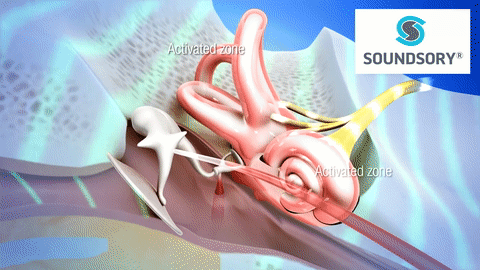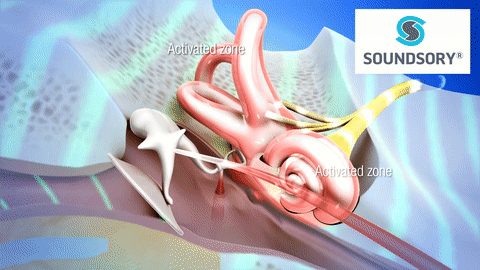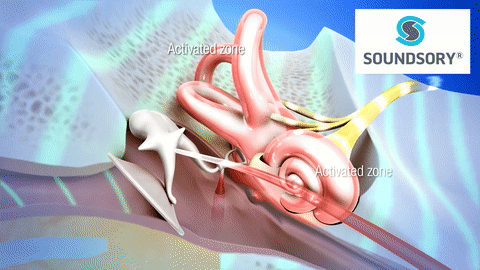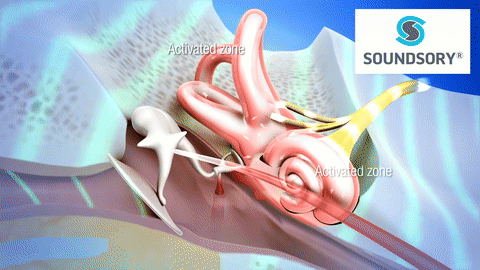
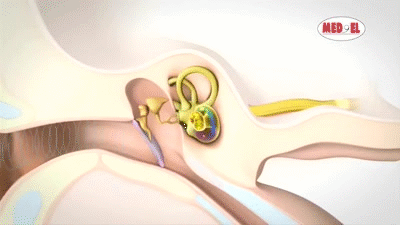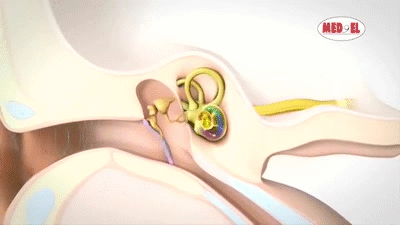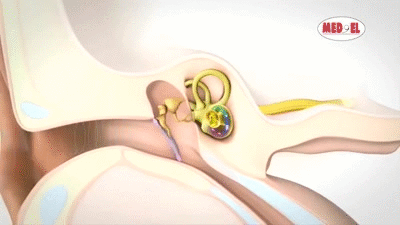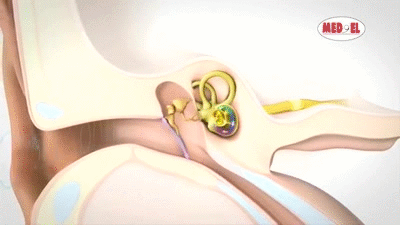
La membrana timpánica (llamada corrientemente tímpano) y los huesecillos, que conducen el sonido desde ella hasta la cóclea (el oído interno) a través del oído medio.
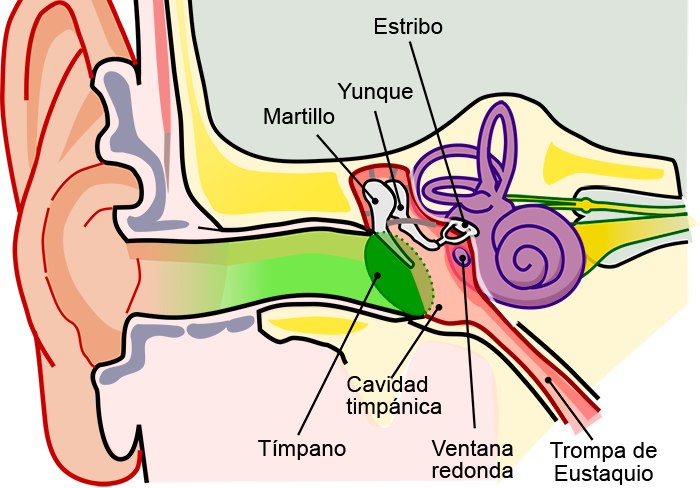
En la membrana timpánica se fija el manubrio o mango del martillo. Este hueso está unido al yunque por unos ligamentos diminutos, por lo que cualquier movimiento del primero arrastra al segundo con él. El extremo opuesto del yunque se articula con la cabeza del estribo y la base de este último descansa sobre el laberinto membranoso de la cóclea en la abertura de la ventana oval.
El extremo final del manubrio del martillo se fija al centro de la membrana timpánica y sobre este punto de inserción tira constantemente el músculo tensor del tímpano, que mantiene tensa dicha estructura. Esta tensión permite que las vibraciones sonoras de cualquier porción de esta membrana se transmitan a los huesecillos, lo que no sucedería si se encontrara relajada. Los huesecillos del oído medio están suspendidos por ligamentos de un modo tal que el martillo y el yunque actúan en combinación como una sola palanca, cuyo fulcro queda aproximadamente en el margen de la membrana timpánica.

La articulación del yunque con el estribo hace que este último: 1) empuje hacia delante la ventana oval y el líquido coclear que está presente al otro lado cada vez que la membrana timpánica se mueve hacia dentro, y 2) tire del líquido hacia atrás cada vez que el martillo se desplaza hacia fuera.
El ajuste de la impedancia entre las ondas sonoras del aire y las del oído coclear está mediado por la cadena de huesecillos
La amplitud de los movimientos de la base del estribo con cada vibración sonora no supone nada más que tres cuartas partes del recorrido que efectúa el manubrio del martillo. Por tanto, el sistema de palanca osicular no aumenta la distancia del desplazamiento del estribo, tal como se cree habitualmente.
Por el contrario, lo que en realidad hace es reducirlo, pero incrementar la fuerza de empuje alrededor de 1,3 veces. Además, la superficie de la membrana timpánica mide un área de unos 55 mm2 . Mientras que la del estribo presenta una media de 3,2 mm2 . Esta diferencia de 17 veces multiplicada por la proporción de 1,3 que corresponde al sistema de palanca hace que la fuerza total a la que está sometido el líquido coclear sea unas 22 veces mayor que la ejercida por las ondas sonoras sobre la membrana timpánica.
Dado que el líquido posee una inercia mucho mayor que el aire, hace falta un grado superior de fuerza para ocasionar la vibración del primero. Así pues, la membrana timpánica y el sistema de huesecillos aportan un ajuste de impedancias entre las ondas sonoras del aire y las vibraciones sonoras en el líquido de la cóclea.
En efecto, el ajuste de impedancias está alrededor del 50 al 75% de la situación ideal para las frecuencias sonoras entre 300 y 3.000 ciclos/s, lo que permite utilizar la mayor parte de la energía portada por las ondas sonoras entrantes.
Si falta el sistema de huesecillos y la membrana timpánica, las ondas sonoras aún pueden viajar directamente a través del aire contenido en el oído medio y entrar en la cóclea por la ventana oval. Sin embargo, en estas circunstancias la sensibilidad auditiva es de 15 a 20 decibelios menor que para la transmisión osicular, lo que equivale a un descenso desde un nivel intermedio de voz hasta otro apenas perceptible

Atenuación del sonido mediante la contracción de los músculos estapedio y tensor del tímpano
Cuando se transmiten sonidos fuertes a través del sistema de huesecillos y desde él al sistema nervioso central, se desencadena un reflejo pasado un período de latencia que solo dura de 40 a 80 ms y que provoca la contracción del músculo estapedio o del estribo y, en menor medida, del músculo tensor del tímpano.
Este último tira del manubrio del martillo hacia dentro mientras que el primero tira del estribo hacia fuera.
Ambas fuerzas se oponen entre sí y de ese modo hacen que el sistema de huesecillos adquiera en su conjunto una mayor rigidez, lo que disminuye mucho la conducción osicular de los sonidos de baja frecuencia, especialmente por debajo de 1.000 ciclos/s.
Este reflejo de atenuación es capaz de reducir la intensidad de transmisión para los sonidos de baja frecuencia de 30 a 40 decibelios, que es más o menos la misma diferencia que existe entre una voz fuerte y un susurro.
Se piensa que este mecanismo cumple una función doble: proteger la cóclea de las vibraciones lesivas ocasionadas por un sonido excesivamente fuerte y ocultar los sonidos de baja frecuencia en un ambiente ruidoso.
La ocultación normalmente elimina un componente importante del ruido de fondo y permite que una persona se concentre en los sonidos por encima de 1.000 ciclos/s, que contienen la mayor parte de la información pertinente para la comunicación vocal.
Otra función de los músculos estapedio y tensor del tímpano consiste en disminuir la sensibilidad auditiva de una persona hacia sus propias palabras. Este efecto es suscitado por unas señales nerviosas colaterales transmitidas hacia estos músculos al mismo tiempo que el cerebro activa el mecanismo de la voz
CÓCLEA
Anatomía funcional de la cóclea
La cóclea es un sistema de tubos en espiral.

Consta de tres tubos enrollados uno junto a otro:
1) la rampa vestibular
2) el conducto coclear o rampa media
3) la rampa timpánica. La rampa vestibular y el conducto coclear están separados por la membrana de Reissner
la rampa timpánica y el conducto coclear están divididos por la membrana o lámina basilar.
Sobre su superficie se encuentra el órgano de Corti, que contiene una serie de células sensibles a estímulos electromecánicos, las células ciliadas. Se trata de los órganos receptores terminales que generan impulsos nerviosos como respuesta a las vibraciones sonoras.

Transmisión de las ondas sonoras en la cóclea: la «onda viajera»
Cuando la base del estribo se desplaza hacia dentro contra la ventana oval, la ventana redonda debe abombarse hacia fuera debido a que la cóclea está encerrada por todas partes por paredes óseas.

El efecto inicial de una onda sonora que llega a la ventana oval consiste en doblar la lámina basilar de la base de la cóclea en dirección hacia la ventana redonda. Sin embargo, la tensión elástica acumulada en las fibras basilares a medida que se curvan hacia la ventana redonda pone en marcha una onda de líquido que «viaja» recorriendo la lámina basilar hacia el helicotrema.

Función del órgano de Corti
- Es el órgano receptor que genera los impulsos nerviosos como respuesta a la vibración de la lámina basilar.
- Los auténticos receptores sensitivos del órgano de Corti son dos tipos especializados de células nerviosas llamadas células ciliadas: una sola fila de células ciliadas internas, que suman unas 3.500 y poseen un diámetro de unos 12 μm, y tres o cuatro filas de células ciliadas externas, que totalizan alrededor de 12.000 y cuyo diámetro no mide nada más que alrededor de 8 micrómetros.
- La base y las caras laterales de las células ciliadas hacen sinapsis con una red de terminaciones nerviosas cocleares.
- Entre el 90 y el 95% de ellas acaban sobre las células ciliadas internas, lo que subraya su importancia especial para la detección del sonido.

La vibración de la membrana basilar excita las células ciliadas
Los cilios diminutos, o estereocilios, llevan un sentido ascendente desde las células ciliadas y entran en contacto o quedan sumergidos en el revestimiento gelatinoso superficial de la membrana tectoria, que se halla por encima de los estereocilios en el conducto coclear.
Estas células ciliadas son semejantes a las que existen en la mácula y en la cresta ampollar del aparato vestibular..
La inclinación de los cilios en un sentido despolariza las células ciliadas, y su inclinación en el sentido opuesto las hiperpolariza. Esto excita a su vez las fibras del nervio coclear que hacen sinapsis en sus bases.
El extremo externo de las células ciliadas está sólidamente anclado a una estructura rígida compuesta por una lámina plana, llamada membrana reticular, sostenida por los pilares de Corti, que están fijos con firmeza a las fibras basilares.
Las fibras basilares, los pilares de Corti y la membrana reticular se desplazan como una sola unidad rígida.

Los potenciales de receptor de las células ciliadas activan las fibras nerviosas auditivas
Los estereocilios son estructuras duras debido a que poseen un armazón rígido de proteínas. Cada célula ciliada posee unos 100 estereocilios sobre su borde apical.
Estos estereocilios van creciendo progresivamente hacia su lado más alejado del modíolo, y la parte superior de los estereocilios más cortos está sujeta por unos filamentos delgados a las porciones posteriores de los estereocilios vecinos más largos.
Por tanto, cada vez que los cilios se inclinen en dirección hacia los más largos, tiran del extremo de los más pequeños hacia fuera desde la superficie de la célula ciliada.
Esto provoca un fenómeno de transducción mecánica que abre de 200 a 300 canales de conducción catiónica, lo que permite el movimiento rápido de iones potasio con carga positiva desde el líquido del conducto coclear adyacente hacia los estereocilios, y esto suscita la despolarización de la membrana de la célula ciliada.
Cuando las fibras basilares se inclinan hacia la rampa vestibular, las células ciliadas se despolarizan, y cuando se mueven en el sentido opuesto se hiperpolarizan, lo que genera así un potencial de receptor alterno en su seno, lo que a su vez estimula las terminaciones del nervio coclear que hacen sinapsis en la base de las células ciliadas.
Se cree que durante la despolarización las células ciliadas liberan un neurotransmisor de acción rápida en estas sinapsis. Es posible que la sustancia transmisora sea glutamato, pero no hay ninguna seguridad al respecto.

Determinación de la frecuencia del sonido: el principio de la «posición»
Los sonidos de baja frecuencia dan lugar a una activación máxima de la lámina basilar cerca de la cúpula de la cóclea, y los de alta frecuencia lo hacen cerca de su base.
Los sonidos de una frecuencia intermedia activan la membrana a una distancia también intermedia entre ambos extremos.
Por añadidura, las fibras nerviosas presentan una organización espacial dentro de la vía coclear, que se conserva durante todo el trayecto desde la cóclea hasta la corteza cerebral.
El registro de señales en los fascículos auditivos del tronco del encéfalo y en los campos receptores auditivos de la corteza cerebral muestra que cada frecuencia sonora específica activa unas neuronas concretas del encéfalo.
Por tanto, el método fundamental empleado por el sistema nervioso para detectar las diversas frecuencias sonoras consiste en determinar el punto más estimulado a lo largo de la lámina basilar, que se denomina principio de la posición para la determinación de la frecuencia sonora.
Se propone que estas frecuencias bajas se diferencian sobre todo por medio del denominado principio de la frecuencia o de la salva.
Los sonidos de frecuencia grave, desde 20 hasta 1.500 a 2.000 ciclos/s, pueden provocar salvas de impulsos nerviosos sincronizados a la misma frecuencia, y estas salvas transmitirse por el nervio coclear hacia los núcleos cocleares del encéfalo.
También se ha planteado que estos últimos núcleos son capaces de distinguir las diversas frecuencias de las salvas.
En realidad, la destrucción de la mitad apical de la cóclea en su integridad, que elimina la zona de la lámina basilar donde se detectan normalmente todos los sonidos de frecuencias inferiores, no suprime por completo la distinción de estos sonidos de frecuencia grave. Determinación del volumen
Determinación del volumen
El sistema auditivo determina el volumen recurriendo a tres procedimientos como mínimo.
- En primer lugar, según sube el volumen sonoro, también aumenta la amplitud de la vibración en la lámina basilar y en las células ciliadas, por lo que estas últimas excitan las terminaciones nerviosas a una frecuencia más rápida.
- En segundo lugar, a medida que aumenta la amplitud de la vibración, hace que se estimule un número cada vez mayor de células ciliadas en la periferia de la porción resonante de la lámina basilar, lo que da lugar a una sumación espacial de los impulsos: es decir, la transmisión a través de muchas fibras nerviosas en vez de solo unas pocas. En tercer lugar, las células ciliadas externas no se estimulan apreciablemente hasta que la vibración de la lámina basilar alcanza una intensidad elevada y la activación de tales células probablemente comunica al sistema nervioso la información de que el sonido es fuerte.
